|
Éléments
d'histoire de la génétique
()
|
retour
accueil, plan du
cours-
|
Les liens renvoient au tableau
historique des sciences.
Les passages en bleu sont
des critiques pédagogiques qui tentent d'essayer de
montrer l'urgence d'une nouvelle formulation de la vie et le
retour à la méthode expérimentale.
Les passages en vert
sont des opinions
personnelles. Les
liens en orange pointent vers des pages plus
détaillées davantage orientées vers
l'analyse des articles originaux.
|
-
|
Sources
La trame initale a été empruntée aux
pages de Alain Bernot et Olivier Alibert sur "La
naissance de la biologie moléculaire" accessibles
à l'adresse http://www.genoscope.cns.fr/externe/HistoireBM/
. De nombreux éléments sont issus des
documents des manuels scolaires de terminale (par exemple
Bordas, collection Tavernier, 1983, et Fernand Nathan,
collection Escalier, 1983, manuels de terminale C et
terminale D). Les principales données de biologie
moléculaire sont essentiellement extraites de:
Biologie moléculaire de la cellule, Alberts et
al., 1995, Médecine-Sciences, Flammarion;
Principes de Biochimie, A. Lehninger, D. Nelson et M.
Cox, 1994, Médecine-Sciences, Flammarion;
Gènes, B. Lewin, 1992,
Médecine-Sciences, Flammarion. Les idées
non-conformistes sur la logique du vivant viennent de: Le
gène et la forme (ou la démythification de
l'ADN), Rosine Chandebois, 1989, Ed. Espaces 34. En 2002
il faut ajouter l'ouvrage Beljanski, un novateur en
biomédecine; concepts, théories,
applications; C.-G. NORDAU et M.S. BELJANSKI, éd.
EVI Liberty Corp, 2001. Enfin, je recommande fortement la
lecture du petit article de Philippe MATHY sur "La
génétique de l'album de famille" dans
Biologie-Géologie (Bulletin de l'APBG), 2-1998,
319-335. Quelques références ont
été prise à l'adresse http://www.ac-amiens.fr/pedagogie/svt/info/hist_genet/index.html.
Des modifications majeures sont en cours en 2006
étant donné mon virage vers la
biologie
théorique. D'autres
données accessibles à tous seront aussi
ajoutées à la suite de la lecture de
L'homme végétal : pour une autonomie du
vivant, Gérard Nissim AMZALLAG, Albin
Michel, 2003 (préface de Bernard Werber) et du
même auteur La raison malmenée. De
l'origine des idées reçues en biologie
moderne, 2002, CNRS Éditions.
On m'a conseillé notamment les ouvrages de Jan Sapp:
Beyond the Gene, New York: Oxford University Press,
1987; Genesis: The Evolution of Biology. New York:
Oxford University Press, 2003 or Where The Truth
Lies. Franz Moewus and the Origins of Molecular Biology.
New York: Cambridge University Press, 1990. (quelques
articles cités sont accessibles sur internet)
Un bref recadrage moderne de l'histoire de la
génétique par Michel Morange, de l'ENS, est
disponible à l'adresse: www.eduscol.education.fr/
D0018/Histoiredelagenetique.pdf
L'histoire de la génétique à Gif sur
Yvette racontée par ses acteurs: http://picardp1.ivry.cnrs.fr/histoire_de_la_genetique.html
Grâce aux nombreux articles classiques,
désormais en ligne (en anglais), les ressources
semblent vraiment immenses, mais il y a d'innombrables
redondances:
http://www.dartmouth.edu/~bio70/papers.html
http://www.esp.org/foundations/genetics/classical/
http://www.bv229.k12.ks.us/biophilia/gsmm.htm
Le fichier http://www.esp.org/foundations/genetics/classical/holdings/b/birth.pdf
contient:
p 1 (9 du fichier pdf) - GREGOR MENDEL'S Letters to Carl
Nägeli. 1866-1873
p 35 (43 du fichier pdf) - De Vries, H.. 1950. Concerning
the law of segregation of hybrids. Genetics, 35(5, pt 2):
30-32
p 39 (47 du fichier pdf)- Correns, C., 1950. G. Mendel's law
concerning the behavior of progeny of varietal hybrids.
Genetics, 35(5, pt 2): 33-41
p 51 (59 du fichier pdf)- Tschermak, E. 1950. Concerning
artificial crossing in Pisum sativum. Genetics, 35(5, pt 2):
42-47
|
-
|
|
|
|
Introduction
|
|
Pour envisager une histoire de la
génétique, on peut s'attacher d'une part
à l'histoire de concepts et d'autre part celle
des acteurs et leur position sociale, ce qui est
assez lié aux applications (sélection
de races il y a un siécle, biotechnologie et
médecine maintenant). Ensuite la
génétique n'a pas de limites si bien
définies que l'on puisse séparer son histoire
de celle de nombreux autres champs scientifiques:
hérédité et embryologie pour n'en citer
que deux.
Lorsque j'ai commencé cette page
en 1998, il y avait encore peu de choses sur internet.
J'avais donc osé l'intituler "une histoire de
la génétique" pour essayer
d'une part de présenter les connaissances que l'on
m'avait transmises lors de ma formation d'enseignant dans un
contexte historique et d'autre part pour montrer les
évolutions actuelles en justifiant ainsi de la
nécessité de réformer le contenu de
l'enseignement.
En 2006, les archives historiques, notamment
américaines, DEVIENNENT ACCESSIBLE À TOUS. On
arrive assez souvent à trouver les articles originaux
(par exemple sur le site http://www.esp.org/).
Mais cela malheureusement ne suffit pas à faire de
l'enseignant (qui, par contre, a souvent une
expérience de la recherche) un historien. C'est
très souvent le contexte social qui est ignoré
et qui ne peut que difficilement être
appréhendé par la seule lecture des articles
scientifiques. On doit donc s'en remettre aux historiens des
sciences. Mais peu de travaux sont accessibles sur internet
à ce jour. De plus il n'est pas rare que l'histoire
soit partisane (comme l'est aussi la présentation
d'un travail scientifique, mais dans ce cas on est mieux
formé pour pouvoir faire la part de ce qui est
opinion et découverte scientifique); en histoire on
est en quelquesorte livré au spécialiste.
C'est donc avec sa propre formation philosophique, ses
convictions humanistes, ses parti-pris, ses ignorances...
que l'enseignant s'efforce de présenter clairement
l'histoire des concepts qu'il enseigne. Qui osera le faire
?
C'est pourquoi, dans cette nouvelle version, je renonce
à écrire UNE histoire et je me contente de
quelques éléments d'histoire qui
renvoient d'ailleurs la plupart du temps à des
pages plus
détaillées
(présentées dans la colonne de gauche) qui
sont les analyses des articles originaux. La tâche est
immense et les éclairages multiples. Cette page, sans
prétendre donc être une histoire à part
entière, est un recadrage, avec mon propre point de
vue forcément partiel, des données historiques
qui s'accumulent dans mes cours et qui expliquent en quelque
sorte ma vision d'ensemble.
|
|
|
|
Plan de cette
page:
|
|
1 - Les
pangènes, particules
héréditaires
2 - Des gènes héréditaires aux
gènes moléculaires
2.1 -Les
gènes héréditaires,
unités mutables appartenant à un groupe de
liaison (le chromosome)
(le gène unité de
mutation et de recombinaison)
2.2 - Du gène bactérien au gène
moléculaire, unité de fonction
()
|
|
pages complémentaires historiques accessibles
TD
corrigé de terminale S: J. Mendel, W.
Johannsen et H. De
Vries
T.
H.
Morgan
transformation
bactérienne (F. Griffith, O. Avery, C.
MacLeod, M.
McCarty...)
G. Beadle
et E. Tatum
|
|
|
|
|
1 - Les
pangènes, particules héréditaires
|
|
J.
Mendel
|
|
En 1866, le moine tchèque,
Johann
Mendel (1822-1884) publie ses
résultats sur des expériences menées
chez le pois (Pisum sativum).
Les travaux de frère Grégor
(Johann Mendel) s'insèrent dans une période de
l'histoire de l'Europe favorable aux sciences qui a vu le
développement de la théorie
cellulaire
(énoncée par Schwann et Schleiden en 1838-1839
et précisée par Virchov en 1858; tout
être vivant est composé uniquement et au moins
d'une cellule; toute cellule est issue d'une autre
cellule). Dans la lignée du travail de nombreux
sélectionneurs de races
végétales et animales, Mendel réalise
un travail que nos contemporains aiment à
présenter comme un modèle du genre
scientifique (hypothético-déductif ou
hypothético-vérificatif), probablement de
façon très peu historique
(http://www.mendelweb.org/MWsapp.html):
un travail basé sur une hypothèse (la
transmission des caractères
héréditaires aux descendants par des
particules présentes chez les parents), prouvé
par la réalisation et l'interprétation
d'expériences rigoureuses de croisements chez
le Pois (Pisum). C'est un des précurseurs de
l'utilisation des statistiques dans l'analyses des
résultats expérimentaux.
|
|
Deux exemples d'analyse de ses
résultats sont donnés dans le TD
corrigé de terminale S: Mendel, Johannsen et De
Vries
|
|
|
|
|
Les travaux de Mendel seront
faussement (http://www.mendelweb.org/MWsapp.html)
"redécouverts" et réinterprétés
par une première génération de
généticiens (comme
Bateson ou De Vries dont un article "Sur la loi de
disjonction des hybrides" est disponible sur
http://www.ac-amiens.fr/pedagogie/svt/info/hist_genet/H_de_Vries.PDF)
qui ont fortement contribué au mythe du
père fondateur. (voir
TD). Ce sont des
expérimentateurs, "discontinuistes" et tout d'abord
non darwiniens, qui s'opposaient alors aux statisticiens
"continuistes" et darwiniens. Hugo
de
Vries (1848-1935)
(travaille sur Oenothera
lamarckiana et) nomme
mutations les changements brusques de
caractères observés dans la descendance. On
sait depuis que cette mutation est due à une
recombinaison par translocation). Il développe la
théorie selon laquelle les mutations président
à l'apparition de nouvelles espèces.
L'histoire n'a pas retenu le terme de
pangène
donné par De Vries aux particules
héréditaires (on peut
rapprocher l'usage de ce terme avec la théorie de la
pangénèse proposée par
Darwin, pour qualifier sa théorie de
l'hérédité des caractères
acquis, courante à l'époque, qu'il faisait
reposer sur l'existence de gemmules; voir par exemple le
cours
de TS).
Mais elle a retenu celui de
gène
donné par le biologiste danois Wilhem
Johannsen
(1857-1927) (ainsi que phénotype et
génotype).
Quelques dizaines d'années plus tard le darwinisme
est incorporé au mendélisme, notamment par
Fisher (1936).
|
|
|
|
|
|
Au début du XXème
siècle on énonce 2 lois EN
HOMMAGE à Mendel:
* 1ère loi: loi de pureté des
gamètes (un hybride formé de deux traits
d'un même caractère donnera en quantités
égales des cellules sexuelles contenant soit l'un
soit l'autre de traits de ce caractère)
(voir
TD).
* 2ème loi: loi de ségrégation
indépendante des caractères (deux
caractères indépendants,
et qualifiés tous les deux de mendéliens, sont
séparés indépendamment dans les
gamètes et réunis indépendamment
à la fécondation; ce qui se voit lors
d'expériences de dihybridisme (voir TD).
Cette loi n'est pas du tout universelle puisque de nombreux
couples de caractères vont avoir une transmission non
indépendante ou liée, ce qui sera la base de
la théorie héréditaire reprise par
Morgan.
|
|
|
|
|
Les "lois de Mendel" (dites
"de John Duff") d'après les manuels
scolaires... on s'éloigne fort de la
vérité
Il me semble
intéressant de proposer les
expériences de Mendel en tant que
données historiques.
Par contre la présentation des "lois de
Mendel" est très criticable car c'est une
réadaptation à des conceptions
modernes (basées sur une
interprétation chromosomique qui est du
domaine de l'hypothèse) de lois empiriques
(établies à partir d'une analyse
statistique qui est un résultat
expérimental qui est toujours valable). On
mélange abusivement deux
conceptions.
|
|
1. Loi d'uniformité
des hybrides de première
génération:
"la première génération
d'hybrides est homogène"
|
tous les hybrides F1 sont
semblables les uns aux autres (même
phénotype et même
génotype);
Pour une caractère donné:
* si les hybrides présentent le
phénotype de l'un des parents, on dit que le
caractère de ce parent est dominant, celui
de l'autre est récessif
* si les hybrides présentent un
phénotype intermédiaireentre ceux des
deux parents, on dot qu'il y a
codominance.
|
|
2. Loi de disjonction (ou
ségrégation) des
caractères en F2:
"les allèles d'un même couple se
disjoignent lors de la formation des
gamètes"
|
les individus F2 sont
différents les uns des autres. Cette
différence s'explique par une disjonction
des caractères allèles au moment de
la formation des gamètes qui sont donc
purs:
* chaque gamète ne contient que l'un ou
l'autre des allèles (loi de pureté
des gamètes)
* les deux catégories de gamètes sont
équiprobables
|
|
3. Loi d'indépendance
des caractères:
|
les phénotypes
observés montrent que la disjonction s'est
faite de manière indépendante pour
les divers couples d'allèles
|
|
Remarque:
cette dernière loi, reprise par des manuels
plus récents (par exemple Terminale C,
Bordas, 1983), s'est transformée en loi de
comportement des allèles soit de
façon indépendante
(phénotypes parentaux et originaux en
même proportion), soit de façon
liée (phénotypes parentaux en
plus grand nombre que les phénotypes
nouveaux)
|
|
Résumé de
l'étape 1
|
Mendel, Johannsen et De
Vries
|
Certains caractères
héréditaires matériels peuvent
être considérés comme
portés par des particules (gènes
ou mieux pangènes pour bien
séparer les notions) transmises aux
descendants par les parents. Ces caractères
matériels sont qualifiés de
caractères présentant une
hérédité mendélienne.
Un changement brusque dans la descendance est une
mutation.
|
retour
plan
|
|
2 - Des
gènes héréditaires aux gènes
moléculaires
|
|
|
2.1 Les gènes
héréditaires, unités mutables
appartenant à un groupe de liaison (le
chromosome)
(le gène unité de
mutation et de recombinaison)
|
|
T.
H. Morgan
|
|
Il semble que ce soit Sutton
(1902) et Boveri (1904) qui proposèrent pour
la première fois (voir des
publications accessibles sur l'ESP (http://www.esp.org/
et des extraits traduits à
l'adresse http://www.ac-amiens.fr/pedagogie/svt/info/hist_genet/index.html)
d'associer les gènes aux chromosomes qui
deviendraient ainsi supports de
l'hérédité (le comportement des
chromosomes lors de la mitose a déjà
été décrit par
Flemming
en 1879 mais c'est vraiment au cours du début de
20ème siècle que se développent les
observations cytologiques).
Un des articles les plus intéressants sur la question
est la revue de Thomas
H. Morgan (1966-1945)
datée de 1910 (Chromosomes and heredity, The
American Naturalist, 44:449-496 (http://www.esp.org/foundations/genetics/classical/holdings/m/thm-10b.pdf;
voir les pages 4 à 13 de la réédition
électronique).
À partir de 1910, Morgan travaille
sur la mouche du vinaigre : Drosophila melanogaster.
Une des premières mutations obtenues par Morgan est
un mâle aux yeux blancs dans une population aux yeux
rouges (caractère sauvage). Comme il n'apparaît
que chez le mâle, l'idée lui vient de
l'associer au sexe (Morgan, T.H. (1910), Sex limited
inheritance in Drosophila, Science, 32, 120,
disponible sur l'ESP : http://www.esp.org/foundations/genetics/classical/thm-10a.pdf;
une traduction partielle en français est disponible :
http://www.ac-amiens.fr/pedagogie/svt/info/hist_genet/Morgan.html).
Une étude est proposée sur la page
annexe sur Morgan.
|
|
Remarque: le terme
de chromosome est difficile à cerner. En
cytologie classique il désigne une structure
colorable qui contient un centromère, zone
d'attachement des fibres kinétochoriales qui assurent
son mouvement.. Le chromosome n'existe donc que sous
forme compacte et lors d'une division. D'autre part, le
chromosome prophasique ou métaphasique ne
possédant qu'un centromère pour deux
chromatides soeurs, le nombre de chromosomes par cellule
lors de ces phases différe de la quantité
d'ADN ou du nombre de copies du génome. Par
extension, chaque nucléofilament a été
appelé chromosome: on parlera alors des chromosomes
déroulés se dupliquant en phase S de
l'interphase. Ainsi certains auteurs (B. Lewin par exemple)
considérent que chaque cellule entrant en mitose
"posséde quatre copies de chaque chromosome"
(Gènes, p23). C'est un point de vue
réductionniste qui tend à considérer la
division cellulaire uniquement au niveau de la transmission
de l'ADN (il est de plus regrettable que les chromosomes
homologues soient qualifiés de copies). Il est de
loin préférable à mon sens pour le
biologiste (et donc pour l'enseignant de SVT) de
privilégier l'aspect cytologique. (Voir par exemple
les pages sur les caryotypes
de l'enseignement de spécialité de terminale
S).
|
|
|
|
La deuxième étape est la
mise en forme de la notion de liaison. Morgan
découvre en effet avec ses collaborateurs d'autres
mutations. Lorsqu'ils étudient la transmission
héréditaire de celles-ci il trouvent qu'elles
forment 4 groupes de liaison: c'est-à-dire
qu'elles ne se répartissent pas au hasard dans les
descendants mais qu'elles présentent une liaison
(linkage, en anglais): elles ont tendance à
rester associées. Plus précisément
Morgan et ses collaborateurs observent 4 groupes de liaison,
c'est-à-dire 4 ensembles de mutations qui ont
tendance à rester associées. Ils
émettent alors l'hypothèse que ces quatre
groupes de liaison sont assimilables aux 4 paires de
chromosomes de la drosophile. La liaison étant
donc "simplement le résultat mécanique de la
localisation des gènes dans les
chromosomes".
|
|
Il semblerait que l'hypothèse de
la liaison entre les gènes ait
été proposée pour la première
fois par Bateson et Punnet en 1905 mais l'idée de
liaison était assortie de celle de répulsion.
C'est Morgan qui proposa l'idée d'une liaison
physique. Ceci est très clairement exposé dans
son bref article du 10 septembre 1911 dans Science (Morgan,
Thomas H. 1911. Random segregation versus coupling in
Mendelian inheritance, Science, 34, 384;
http://www.esp.org/foundations/genetics/classical/holdings/m/thm-1911a.pdf).
« Instead of random segregation in Mendel's sense we
find "associations of factors" that are located near
together in the chromosomes. Cytology furnishes the
mechanism that the experimental evidence demands.»
(Au lieu d'une ségrégation aléatoire au
sens de Mendel, nous trouvons des "associations de facteurs"
qui sont situés sur les chromosomes. La cytologie
nous fournit le mécanisme que les résultats
expérimentaux demandent).
|
|
Morgan, à partir d'environ 400
gènes connus chez la drosophile, retrouva toujours
les 4 groupes de liaisons. Depuis la théorie des
groupes de liaison n'a cessé de se confirmer. Voici
quelques organismes, avec le nombre de groupes de liaison
trouvé chez chacun, à comparer avec leur
nombre de chromosomes :
|
organisme
|
nombre de groupes de
liaison
|
nombre de paires de
chromosomes (n)
|
|
Drosophile
|
4
|
4
|
|
Pois de
senteur
|
7
|
7
|
|
Pois
|
7
|
7
|
|
Muflier
|
8
|
8
|
|
Maïs
|
10
|
10
|
|
|
La part du mystère ou Pour aller
au-delà de la notion de groupe de liaison
dans une perspective moderne (de biologie
théorique dont un aperçu peut être vu
dans une page
sur les modèles de René
Thom)
Il faut bien se garder de
considérer qu'avec l'interprétation
strictement chimique de l'hérédité et
du rôle des chromosomes on a compris
l'hérédité. Les chromosomes sont des
organites d'une cellule vivante en division. Les
chromosomes ne sont que les organites visibles mais les
fonctions (les dynamiques métaboliques) restent
cachées.
Pour exprimer ces
notions encore peu formalisées je suis amené
à utiliser un vocabulaire un peu abscons mais qui
sera progressivement clarifié (voir les
pages
sur le formalisme
Thomien).
La forme métabolique héritable
(définie dans l'espace de régulation avec le
temps comme variable supplémentaire) est
inaccessible.
Chaque groupe de liaison correspond à une
unité métabolique (dynamique) dont la trace
matérielle est le chromosome. Le chromosome est
l'organite de la division cellulaire et, donc une forme
(dynamique) de la transmission héréditaire de
cellule à cellule ou d'organisme pluricellulaire
à cellule sexuelle.
Une liaison partielle à un chromosome peut rendre
compte de redondances métaboliques ou d'interactions
entre chacune des unités métaboliques
associées à chaque chromosome. Une
recombinaison chromosomique peut aussi résulter de
composition entre unités métaboliques.
|
|
|
|
La dernière étape concerne
l'hypothèse de la disposition linéaire des
gènes sur le chromosome. Cette idée est
née la notion de liaison partielle obtenue pour
certains gènes. En effet certains gènes,
appartenant au même groupe de liaison (et donc au
même chromosome d'après ce que l'on a
supposé), se séparent tout de même
à la méïose mais pas toujours. Pour
mesurer le degré de liaison on réalise des
croisements tests ou test-cross par exemple entre un hybride
F1 et une double récessif parental.
|
|
|
Un test-cross ou
croisement test
|
|
On considère deux
mutations conduisant à des phénotypes
récessifs:
corps noir (b pour black) et ailes
vestigiales (vg); les allèles
sauvages sont notés + (corps gris,
ailes longues)
|
|
femelle hybride (F1) ++
// b vg X b vg // b vg mâle double
récessif (parental: F0)
|
|
+
+
|
b
vg
|
+
vg
|
b
+
|
|
b
vg
|
+ + // b
vg
|
b vg // b
vg
|
+ vg // b
vg
|
b + // b
vg
|
phénotypes
et
pourcentages
|
phénotypes
parentaux
|
phénotypes
recombinés
|
corps gris,
ailes longues
41,5%
|
corps noir,
ailes vestigiales
41,5%
|
corps gris,
ailes vestigiales
8,5%
|
corps noir,
ailes longues
8,5%
|
L'interprétation d'un test-cross se fait en
terme de recombinaison. c'est-à-dire de
liaison partielle: les allèles b et
vg sont partiellement liés (dans 83% des
cas). Ils se séparent (se recombinent
entre eux dans les gamètes) pour 17% (8,5%
+8,5%) des cas. On dit que le taux de recombinaison
est de 17%.
|
|
|
|
|
L'interprétation cytologique de la
liaison partielle a été proposée par
Morgan comme étant un crossing-over, ou
enjambement. Les chromatides échangeraient des
segments lors de la méïose. Le
mécanisme du crossing-over est loin d'être
élucidé . Etant donné le but de ces
pages je vous laisse vous reporter à des ouvrages
spécialisés (Gènes par exemple, ch.
26).
Quelques remarques:
1 - Un taux de 50% de recombinaison ne peut en principe pas
être atteint car cela correspond à deux
gènes indépendants statistiquement. Il ne faut
pas oublier que la notion de gènes liés est
une notion statistique. Vu de cette manière, le
chromosome est un groupe de liaison.
2 - Les crossing-over ne peuvent pas être vus, si ils
existent, lorsqu'ils affectent deux chromatides d'une
même chromosome ou lorsqu'ils affectent des
gènes homozygotes (c'est-à-dire
présentant les mêmes allèles sur la
paire d'homologues d'un individu).
3 - Pourquoi le mâle de drosophile
présente-t-il une liaison absolue des ses
gènes alors qu'il semble que la liaison partielle
soit la règle et est la plupart du temps identique
chez les deux sexes ?
|
|
|
|
|
On en arrive à la dernière
hypothèse de Morgan et des ses collaborateurs: les
crossing-over ont autant de chance de se produire en tout
point du chromosome: le taux de recombinaison
(c'est-à-dire la fréquence des crossing-over
dans leur modèle) entre deux gènes est
indépendant de la nature de chaque gène, il ne
dépend que de leur position relative sur le
chomosome, ce qui revient à dire que le crossing-over
est un phénomène mécanique (on peut
aussi le présenter comme étant le fruit du
hasard mais cela ne veut pas dire grand chose car les
déterminismes mécaniques ne sont pas le
hasard, qui, pour de nombreux biologistes, n'est que la
mesure de notre ignorance). Ainsi plus deux gènes
sont éloignés plus le taux de recombinaison
sera élevé. C'est la notion de distance
génétique. Le taux de recombinaison,
c'est-à-dire la fréquence des crossing-over
entre deux gènes liés, est d'autant plus
élevé que les loci (le locus étant le
site d'un gène) de ces gènes sont
éloignés physiquement sur le chromosome.
Morgan et ses collaborateurs établissent ainsi une
relation entre un résultat statistique (la liaison
partielle) et une distance physique. Elle leur permet
d'établir des cartes factorielles qui sont la
schématisation des positions respectives des
gènes évaluées à partir des
pourcentages de recombinaisons. Morgan présente avec
Alfred
Sturtevant (1891-1970) en 1913 la
première carte génétique
(voir extrait de l'article à
l'adresse: http://www.ac-amiens.fr/pedagogie/svt/info/hist_genet/Sturtevant.html).
Depuis on a défini le centiMorgan (cM) comme
unité de recombinaison: 1 cM étant égal
à 1% de crossing-over.
Comme pour Mendel, on peut souligner la
chance de Morgan. La chance vient du choix de la Drosophile
qui ne présente pas de crossing-over chez le
mâle (liaison absolue entre deux allèles, soit
un taux de recombinaison nulle) alors qu'il existe chez la
femelle (liaison partielle soit un taux de recombinaison
différent de zéro). Cette chance est bien
sûr une limite quand il faut
généraliser.
|
|
Voici quelques éléments
d'une carte chromosomique ou factorielle très
simplifiée de la drosophile femelle (les chiffres
indiquent des distances en cM) (il semblerait que l'on ne
connaisse pas de gènes positionnés avec
certitude sur le chromosome Y !!!):
chromosome n°1 (X): 2 - w: oeil blanc (+
oeil rouge vif); 6 - ec : echinus: oeil rugeux (+
oeil lisse) ; 13 - cv : cross veinless: nervures
transversales absentes (+ nervures transversales
présentes); 43 - s : sable: corps noir (+
corps gris)- centromère
chromosme 2: 48 - b : black : corps noir (+
corps gris) ; pr : purple : oeil pourpre (+ oeil
rouge vif); centromère ; 67 - vg : ailes
vestigiales (+ ailes longues)
chromosome 3: -----(bras long)-----;
centromère; 70 - eb : ebony: corps noir (+
corps gris)
chromosome 4: punctiforme: pas de recombinaison
observées
Quelques références:
Morgan, T.H. (1911) The origin of nine wing mutations in
Drosophila, Science, 33, 496
Morgan, T.H. (1911) The origin of five mutations in eye
color in Drosophila and their mode of inheritance, Science,
33, 534
Morgan, T.H., Sturtevant, A.H., Muller, H.J. and Bridges, C.
(1915) The mechanism of mendelian heredity;
New-York.
|
|
|
|
Un insecte
très particulier: la
drosophile
ou mouche du vinaigre (Drosophila melanogaster)
bon ou mauvais "outil" ?... les deux très
certainement
|
|
|
|
Les raisons de son succès dans
l'analyse génétique:
- sa taille: inférieure au
millimétre, on peut en élever plusieurs
centaines dans un flacon en les nourissant à peu
de frais de levures ; mais pas trop petite, ce qui permet
les observations morphologiques directement sous la loupe
binoculaire après les avoir endormies à
l'éther; plus récemment la taille des
larves est suffisante pour permettre des marquages
isotopiques ou fluorescents facilement localisables sur
des coupes, même en microscopie optique (ce qui est
beaucoup plus simple que l'utilisation d'un microscope
électronique à transmission comme cela est
nécessaire pour les bactéries).
- la briéveté de son
cycle de vie (une dizaine de jours) et sa
prolificité (200 à 300 oeufs par femelle
pour l'ensemble de sa vie): on peut donc observer environ
30 générations par an à
température ambiante.
- la facilité d'obtention
d'individus mutés présentant des mutation
stables : Morgan a du élever de très
nombreuses générations avant d'obtenir son
mutant mâle à yeux blancs. A partir de 1926
Muller propose l'utilisation de rayons X qui par
irradiation des adultes augmente considérablement
l'apparition de phénotypes mutants stables. On
connait actuellement de très nombreuses mutations
stables de la drosophile (j'ai lu le chiffre de 400 mais
il date certainement un peu...).
|
|
MAIS... cela ne doit pas faitre oublier
certaines spécificités:
- le nombre de chromosomes est
très petit (4 paires) avec un déterminisme
chromosomique du sexe très simple (XX chez la
femelle et XY chez le mâle); la compaction du
génome peut peut-être expliquer certaines
caractéristiques héréditaires non
généralisables...
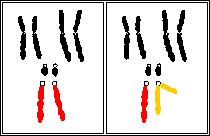
schémas théoriques de caryotypes de
drosophile femelle à gauche et mâle à
droite;
les cercles représentent les centromères;
2n = 8;
3 paires d'autosomes dont une paire punctiforme et une
paire d'hétérochromosomes
(gonosomes)
- certaines cellules larvaires en
interphase comme les cellules des glandes salivaires (que
les élèves connaissent bien pour les
observer chez la larve de chironome, un autre petit
Diptère) possédent des chromosomes
géants ou chromosomes polyténiques
ou polytènes (le degré de
polyténie étant le nombre de copies de
chaque chromatide contenue dans un chromosome
géant). Ils sont moins condensés que les
chromosomes classiques. Leur taille est due à une
duplication des chromatides en plusieurs milliers
d'exemplaires (5.000 environ chez la drosophile). Il ne
semble pas que cette replication empêche la
transcription des gènes actifs, cet état
polytène étant un état
déroulé (ce qui permet de fixer des sondes
radioactives par hybridation et de localiser certains
gènes). En fait on pense maintenant que les zones
transcrites forment des boursouflures ou puffs (anneaux
de Balbiani) mais il y a des exceptions. Ces chromosomes
polyténiques peuvent être colorés
aisément par des colorants basiques comme le vert
de méthyl et l'on observe alors une alternance de
bandes transversales claires et sombres
(chomomères). Il y a donc une structure
répétitive présentant une
affinité particulière pour le colorant. On
ne connaît pas, comme dans le cas des colorations
des caryotypes, les causes moléculaires de cette
affinité. Il faut noter que dans le cas de
mutations (Painter semble être le premier à
l'avoir montré dans les années 1930), on
observe des altérations morphologiques des
colorations des bandes des chromosomes
polyténiques. Il a même été
possible de relier certaines mutations avec
l'altération d'une bande spécifique. On a
donc une carte cytologique des chromosomes. Chaque
bande correspond à environ 25.000 paires de bases
et pour l'instant on ne connaît pas la relation
qu'il existe entre une bande et le nombre d'unités
de transcriptions qui sont probablement plusieurs
(à lire Gènes, ch. 25).
- le développement embryonnaire
est très complexe comme chez de nombreux insectes
et se passe par paliers (métamorphoses): on parle
de développement indirect. L'oeuf, pondu par la
femelle (qui le féconde au passage dans son
oviducte grâce aux spermatozoïdes qu'elle a
stockés lors d'un accouplement), donne naissance
à une larve apode (sans pattes), vermiforme
(asticot) qui se nourrit activement sans l'aide des
parents et croît rapidement sans stades
différenciés. Au bout de quelques jours la
larve s'enpupe, c'est-à-dire se
métamorphose en pupe, un stade larvaire immobile
où la larve, incluse dans un membrane
desséchée de couleur sombre, ne se nourrit
plus et subit de nombreuses transformations
morphologiques, anatomiques et physiologiques: c'est la
mue imaginale qui donne l'adulte. En effet l'adulte, au
bout de quelques jours à température
ambiante sort de la pupe. C'est un adulte ailé,
avec 3 paires de pattes et qui a déjà
atteint quasiment la maturité sexuelle,
d'où la briéveté du cycle (voir
détails sur la page
sur le
développement).
Il est essentiel de noter que du fait des
caractéristiques de ce développement
embryonnaire (que l'on qualifie d'adapté au milieu
aérien) les expériences de manipulation
(d'embryologie expérimentale) sont impossibles
à réaliser sur la larve ou la pupe. En
effet, toute rupture de la cuticule entraîne une
perte du précieux liquide protecteur et donc
provoque la mort de l'embryon. Comme on ne peut pas
déplacer les lignées cellulaires
embryonnaires, toutes les conclusions faites sur le
fonctionnement des différents gènes mis en
évidence chez la drosophile ne peuvent pas prendre
en compte le rôle des cellules environnantes
puisqu'on ne peut pas tester expérimentalement
leur action. Ainsi, d'un bon "outil"
génétique, la drosophile est devenu par
commodité mais indûment un mauvais "outil"
de la biologie du développement. Il est toujours
dangereux de prendre une seul espèce comme
modèle expérimental (ces idées me
semblent particulièrement importantes et viennent
de Rosine Chandebois dans son livre: "Pour en finir avec
le darwinisme", p 69).
Une brève de La Recherche (L'inconnu du
placenta, Olivier Donnars, mai 2003, 364, p 18) met
l'accent sur le lien souvent indu fait entre un
gène non fonctionnel et une anomalie de
développement ou une mort
prématurée: ce n'est pas parceque la
modification d'un gène cause la mort de l'embryon
que l'on sait OU et QUAND (au niveau cellulaire) ce
gène non fonctionnel a été la cause
de ce qui a conduit à la mort de l'embryon. Dans
le cas de la
génétique
du développement en milieu
fermé comme on pourrait
appeller les expériences sur la drosophile, on
s'expose à ne jamais pouvoir comprendre OU et
QUAND les pseudo-gènes du développement ont
été exprimés.
|
|
|
|
|
|
|
|
Quels sont les points communs entre la
carte factorielle (appelée aussi
génétique ou encore chromosomique),
établie par des méthodes statistiques, et la
carte cytologique établie par hybridation de
sondes radioactives ou par observation de modification de
bandes colorées après mutation ?
|
|
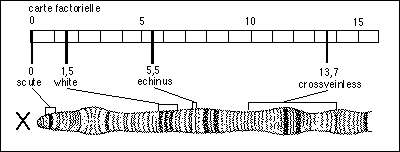
Un exemple très
simplifié de correspondance entre la carte
factorielle et la carte cytologique
établie à partir des bandes colorées
sur le chromosome X géant de la drosophile (Nathan,
TC, TD, 1983)
|
|
|
|
|
|
Globalement, la correspondance est bonne
si l'on s'en tient à la disposition linéaire
des gènes. Par contre les distances sont nettement
différentes. mais la carte cytologique n'est pas
beaucoup plus précise que la carte factorielle. On ne
connaît pas le degré de compaction de l'ADN
dans les chromosomes polytènes... En tout cas on ne
devrait employer le terme de carte chromosomique que
lorsque l'on tient compte à la fois des
données statistiques et cytologiques.
|
|
|
|
|
Résumé de
l'étape 2
|
Sutton et Boveri, Morgan et
Sturtevant
|
Certains caractères
matériels sont liés
héréditairement (sont transmis avec)
avec les chromosomes. Les chromosomes peuvent donc
être considérés comme le
support matériel de
l'hérédité mendélienne.
Certains caractères matériels ne sont
liés que partiellement aux chromosomes (lors
de la transmission ils changent de chromosome).
Cette liaison partielle peut être
interprétée en termes de
recombinaison ou crossing-over (échanges de
fragments entre chromosomes). La mesure de la
fréquence des ces recombinaisons de
façon statistique est
interprétée en termes de distance
entre les gènes disposés
linéairement sur le chromosomes, le
crossing-over ayant autant de chance de se produire
en tout point du chromosome. Ces distances
permettent d'établir une carte
factorielle (ou statistique ou
génétique ou chromosomique s.s.) des
gènes chromosomiques portant les
caractères matériels transmis selon
les lois énoncées
ci-dessus.
Certaines mutations ont aussi
un support chromosomique qui peut être mis en
évidence par les techniques de colorations
de bandes des chromosomes. On établit ainsi
une carte cytologique des gènes
associés aux mutations.
Les gènes sont donc
alors des unités de recombinaison et de
mutation : un gène est une unité
mutable appartenant à un groupe de liaison
(statistique) : le chromosome.
La plupart des
caractères matériels peuvent
être associés à un ou plusieurs
gènes chromosomiques. Pangènes et
gènes sont des notions qui se recoupent mais
ne se superposent pas. On peut parler d'un
modèle héréditaire
chromosomique.
|
retour au
plan de cette page
|
|
|
|
|
|
|
|
|
|
|
|
Remarques:
Les limites de l'analyse statistique mendélienne sont
assez difficiles à cerner mais on peut s'y essayer de
façon très imparfaite:
* la notion de lignée pure statistique
est dépendante d'une observation
phénotypique et, étant donné que le
lien n'est pas absolu entre génotype et
phénotype (vois aussi cours de terminale S), la
notion de lignée pure n'est pas neutre, elle repose
sur une observation statistique dont on ne connaît pas
la causalité matérielle permanente (si elle
existe). Il était d'usage de rappeler dans les
manuels scolaires des éditions
précédentes combien cette relation
était complexe; on citait par exemple les animaux
albinos (souris, rats...) chez qui l'albinisme était
relié à des caractères comportementaux
(agressivité...), ou encore chez la drosophile la
découverte de plusieurs gènes
différents codant pour la couleur sombre du corps
(sable, ebony, black...) ; ou enfin la longueur des oreilles
chez certaines races de lapins que l'on sait être sous
la dépendance de plusieurs gènes additionnant
leurs effets.
La liaison
gène-caractère
Ces notions sont
extraites de l'article de Philippe MATHY sur la
génétique de l'album de famille puis
complétées à ma
façon
|
|
caractère
mendélien s.s. = caractère
monogénique
|
correspondance biunivoque
entre génotype et phénotype (un
gène - un caractère), les
caractères et les gènes étant
considérés comme
indépendants
ces caractères sont extrêmement rares
chez les plantes et les animaux, leur existence est
à démontrer chez l'homme.
|
|
caractère
mendélien élargi (s.l.)
par exemple au sens des
caractères obéissant aux lois de John
Duff
|
polygénie
|
des gènes - un
caractère ; tout le monde s'accorde à
dire que pour l'homme la plupart des traits
phénotypiques sont conditionnés par
plusieurs gènes
|
|
pléiotropie
|
un gène - des
caractères ; un même gène peut
conditionner plusieurs caractères
|
|
épistasie
|
interaction non
alléliques entre gènes : des
gènes conditionnant des caractères
différents pouvant moduler (gènes
"modificateurs") l'aspect de l'un des
caractères
|
|
les effets cités entre
gènes ou entre caractères sont
additifs et des interactions peuvent avoir lieu
mais la correspondance gène-caractère
n'est pas remis en cause : le gène
associé au caractère mendélien
au sens large est une particule transmissible
héréditairement, il a un support
matériel partagé par les parents et
transmis aux descendants
|
|
Pour éviter une
glissement de sens il suffit de définir
précisément la notion de
caractère ; en voici un essai en utilisant
le terme matériel au sens de matière
(objet des sens et quantifiable
expérimentalement) :
Un caractère
matériel peut être
défini comme un trait phénotypique
matériel de l'individu, mesurable
expérimentalement à l'aide de
paramètres physiques et susceptible
d'être décomposé en
éléments moléculaires. On peut
convenir d'appeller gène ce qui est
transmissible héréditairement
et qui est associé à un ou
plusieurs caractères et de façon plus
ou moins univoque. Il est aussi essentiel de
souligner que si le caractère est
associé au gène, ce n'est pas le
gène qui détermine ou gouverne
l'apparition du caractère mais la cellule
qui exprime ou non le gène. Le gène
est la matière nécessaire mais pas
suffisante à l'expression d'un
caractère. Les caractères
matériels d'un adulte ne sont pas
portés par le gamète même si le
gamète comporte les gènes
nécessaires à l'expression de ce
caractère qui pourra ou non être
exprimé. Le gène est ici une
particule héréditaire, une
unité de transmission d'un caractère
matériel.
Il est donc inadéquat de
parler du caractère "couleur des yeux" si
l'on se réfère à autre chose
qu'à l'origine matérielle de la
pigmentation de l'iris. Le déterminisme
(causalité) d'un caractère
matériel est donc sous le contrôle
d'éléments matériels dont les
gènes (au sens de portions d'ADN) sont
certainement un des éléments
essentiels. A ce sujet il me paraît tout
à fait souhaitable, comme le propose
Philippe MATHY, de remplacer les termes "gouverne"
ou "contrôle" par "est associé
à". Ainsi on pourra dire que chez l'homme le
caractère matériel couleur des yeux
est certainement associé à plusieurs
gènes dont certains sont déjà
connus. On peut aussi critiquer le fait d'attribuer
toutes les couleurs des yeux à un
caractère unique : si un caractère
matériel comme yeux bleus peut être
relativement clairement défini il est tout
à fait possible que la couleur jaune d'un
oeil ne soit plus du tout due à un
problème de pigmentation mais à un
problème par exemple de vascularisation. Il
faut donc se méfier et essayer de
déterminer les caractères
matériels au plus près des
molécules. Si cela est assez facile pour un
procaryote, cela devient nettement plus difficile
lorsque l'on s'intéresse à des
eucaryotes et a fortiori à l'homme. De la
même façon il est tout à fait
légitime de parler du caractère
"hémoglobine normale" pour un homme. Par
contre le caractère "sain" ne veut rien
dire, ni le caractère "agressif" ou
"homosexuel". Le mot caractère, pris au sens
de caractère matériel, ne s'applique
pas aux composantes comportementales d'un individu,
encore moins d'un homme. Toujours dans ce sens une
maladie humaine est un événement qui
n'est pas réductible à l'ensemble de
ces caractères matériels
associés à des symptômes (et
donc aux gènes associés à ces
caractères matériels) même
s'ils constituent une voie de recherche.
Remarques :
*Pour éviter de diluer la causalité
d'un caractère physiologique ou même
comportemental d'un organisme on pourrait essayer
de formaliser les relations entre un
caractère et les composantes du vivant :
matière, énergie et information. Un
caractère biologique au sens large comme par
exemple la mobilité d'une bactérie
(pour prendre un exemple peut-être plus
simple) est sous la dépendance de
déterminismes matériels,
énergétiques et informatifs. Par
exemple pour Escherichia coli : la mobilité
fait appel à des flagelles dont la structure
est connue, le mécanisme de rotation et son
déterminisme énergétique a
même été assez clairement
précisé. On connaît de
façon séparée les gènes
associés à chaque caractère
matériel comme les protéines
flagellaires, les molécules d'ATP
synthase... Du point de vue éthologique, le
chimiotactisme a aussi été mis en
évidence et quantifié et on imagine
aisément un support moléculaire avec
des chimiorécepteurs... Mais que sait-on de
l'information qui détermine un comportement
et est-il totalement déterminé ?
Actuellement la biologie moléculaire
matérialiste propose une information
codée par l'ADN, le comportement
résultant du fonctionnement imbriqué
des différentes unités du vivant
matériel. le déterminisme est total.
Tout comportement étant alors
obligatoirement le résultat des composantes
matérielles génétiques et
environnementales. Le hasard n'est que le flou de
la pensée (ou du programme informatique) qui
ne peut arriver à imaginer (ou à
modéliser et donc à calculer)
l'interaction simultanée de causes aussi
innombrables. Il n'y a qu'une illusion de
comportement.
* Voir le nouveau
(2003) cours de 1ère
S pour un essai de
clarification des termes de
"caractère", "génotype"
et "phénotype".
|
* tant que l'on considère un, deux, voire trois
caractères, l'analyse statistique est possible mais
dès que l'on atteint des cas de polyhybridisme,
l'analyse statistique expérimentale n'est
plus possible: réaliser 16 384 croisements
(128x128) pour tester les 128 génotypes
théoriques possibles (27) que l'on peut
imaginer avec 7 couples de caractères n'est pas
réaliste.
* de la même manière, l'analyse
statistique des F3 qui suppose de nombreux
croisements est pratiquement impossible
au-delà de deux caractères.
* enfin l'application à l'étude de
l'hérédité humaine
me paraît indue. Il ne peut y avoir
d'analyse statistique de la descendance pour des familles
qui dans le meilleurs des cas atteignent la dizaine
d'enfants. Que l'on fasse une analyse
héréditaire à l'aide de la
méthode préalablement justifiée se
conçoit mais l'étude des arbres
généalogiques humains ne permet en aucun cas
de trouver une justification de l'analyse statistique de la
transmission des caractères (voir
cours
de TS).
Quel est donc
l'intérêt pédagogique
d'une étude mendélienne ? Un
intérêt historique certain. Aussi bien pour
l'histoire des concepts et des acteurs que pour celle de la
méthode expérimentale en
précisant bien d'une part l'hypothèse
principale de Mendel (particules transmissibles support de
l"hérédité) et d'autre part en
présentant les résultats de croisements et
l'analyse de ces résultats sans autre données
que celles de Mendel. Viendra ensuite, après les
observations cytologiques et chromosomiques,
l'hypothèse des chromosomes, support de
l'hérédité (notion très
imparfaite qui n'est qu'une étape dans la
démarche). Mais il est ridicule de vouloir illustrer
les mécanismes cytologiques et chromosomiques de la
méiose et de la fécondation à l'aide de
ces analyses statistiques. Si l'on se réfère
au programme de terminale S actuel c'est pourtant cette
démarche qui est adoptée. On fait observer les
phénomènes cytologiques et chromosomiques
intervenant lors d'un cycle de reproduction. Puis on
présente l'analyse statistique comme une
justification des mécanismes chromosomiques. La
démarche est inversée. Il est indispensable de
la rectifier. (Pour un lecteur attentif de ces pages vous
noterez que le problème de fond se résume
d'une part à l'abandon de la méthode
expérimentale, d'autre part à une conception
erronnée du vivant, la vie étant
réduite à des
mécanismes).
Jusqu'ici la
génétique et l'embryologie peuvent être
concilées. Mais c'est avec le développement de
la biologie moléculaire que la notion de gène
va échapper aux théoriciens de
l'hérédité. Certains auteurs (comme
Carlson, ), en ce début du XXIème
siècle considèrent que l'on peut garder et
approfondir le lien entre hérédité et
chromosomes et s'affranchir de la vision moléculaire
devenue obscure.
retour
plan
|
|
|
|
|
|
2.2 Du
gène bactérien au gène
moléculaire, unité de fonction
|
|
|
transformation
bactérienne (F.
Griffith)
G. Beadle
et E. Tatum
|
|
Les gènes mendéliens et
morganiens sont chromosomiques et donc eucaryotes. Mais
dès les années 1930 les microbiologistes
s'efforcent de prouver que la notion de gène est
aussi pertinente pour la transmission des caractères
héréditaires des bactéries. C'est
probablement un point fondamental qui va faire basculer la
génétique vers la génétique
moléculaire en se séparant de la cytologie,
quasiement inaccessible pour de si petits
organismes.
C'est
Fred Griffith (1877-1941), en
1928, qui découvre la transformation
bactérienne, mais s'il prouve la
présence d'un facteur transformant, il ne le met pas
en relation avec l'ADN, ce qui ne sera fait que 10 ans plus
tard par Oswald Avery et ses collègues. De plus ces
tranbsformations ne SONT PAS RELIÉES AUX MUTATIONS et
donc aux gènes des eucaryotes.
Expérience de
Fred Griffith (1928)
avec commentaires à partir du vocabulaire et
des notions actuels
|
|
Il utilise deux souches de
pneumocoques. La bactérie Streptococcus
pneumoniae (ou Pneumococcus), est virulente
grâce notamment à une capsule
polysaccharidique qui la protège de la lyse
lors de la phagocytose. On parle de forme S
(smooth = lisse en anglais) car la colonie
sur gélose prend un aspect lisse et
brillant. Il apparaît dans les cultures des
souches R par mutation qui perdent leur
capsule. Les colonies de souches R ont un aspect
rugueux et mat (rough = rugueux en anglais).
Il existe plusieurs souches nommées SI-RI,
SII-RII, SIII-RIII suivant les types de
polysaccharides de la capsule. Il existe des
mutations reverse mais toujours au sein d'une
même souche (RII vers SII par
exemple).
Griffith, en 1928, inocule
à une souris une petite quantité de
pneumocoques RII avec une grande quantité de
pneumocoques SIII "tués" à 60°C
pendant 30 min. Le sang prélevé sur
des souris sacrifiées ultérieurement
est mis en culture et il apparaît des
bactéries de forme SIII vivantes et
virulentes (avec une capsule).

Griffith suggère que le
SIII tués ont fourni une protéine
spécifique aux RII leur permettant de
fabriquer une capsule. Il existe donc une
transformation de la souche RII en souche SIII
à moins de proposer une reviviscence des
SIII. Ce principe transformant reste
à découvrir.
Quelques points d'analyse de
cette expérience:
- une mutation reverse aurait
donné des SII
- on pourrait imaginer que les
RII se glissent dans les capsules des SIII, mais
si l'on réalise l'expérience avec
des SIII débarassées de leur
capsule on obtient toujours des SIII vivantes
avec une capsule
- le traitement à
60°C est suffisant pour dénaturer
les protéines (de façon
irréversible): jamais aucune souche
"tuée" ne devient virulente, sauf par
mutation...
|
Ayant trouvé
sur internet la publication originale de Griffith, mon
analyse a quelque peu changé: voir une
page
complémentaire
L'article de Luria, S. E., and M.
Delbrück de 1943 : Mutations of bacteria from virus
sensitivity to virus resistance (Genetics, 28: 491-511
-http://www.esp.org/foundations/genetics/classical/holdings/l/slmd-43.pdf)
est considéré par certains comme le premier
papier qui expose clairement des arguments (statistiques)
tendant à prouver que les bactéries ont aussi
des gènes (raisonnement basé sur l'apparition
de mutations). «We consider the above results as
proof that in our case the resistance to virus is due to a
heritable change of the bacterial cell which occurs
independently of the action of the virus.»
(Nous considérons que les
résultats ci-dessus sont la preuve que, dans notre
cas expérimental, la résistance aux virus est
due à l'apparition d'un caractère
héréditable chez la cellule
bactérienne, de façon indépendante de
l'action du virus).
Lorsque George Beadle et Edward Tatum en
1941 cherchent un matériel expérimental pour
établir un lien entre mutants et métabolisme,
il choississent Neurospora crassa, un champignon, et
donc un eucaryote, car, malgré le fait qu'Edward
Tatum soit microbiologiste, la notion de gène
bactérien n'est pas encore admise
(voir page sur
ces deux auteurs). Par contre
lorsque Tatum travaillera seul après 1946 il
retournera aux bactéries pour lesquelles la notion de
gène étaient désormais
admise.
retour
plan
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
http://www.dartmouth.edu/~bio70/papers.html
Scott Gilbert, The
Embryological Orgins of the Gene Theory (http://zygote.swarthmore.edu/gene1.html)
Scott Gilbert, Cellular
Politics: Ernest Everett Just, Richard B. Goldschmidt, and the
Attempt to Reconcile Embryology and Genetics (http://zygote.swarthmore.edu/gene2.html)
Scott Gilbert, Induction and
the Origins of Developmental Genetics (http://zygote.swarthmore.edu/gene3a.html)
Scott Gilbert, Enzymatic
Adaptation and the Entrance of Molecular Biology into Embryology
(http://zygote.swarthmore.edu/gene5a.html)
De
l'information génétique au programme
génétique
La deuxième étape pourrait
être qualifiée de période de domination des
biochimistes et des physiciens. Elle correspond au passage de la
notion de gène "héréditaire", particule
chromosomique et support des caractères
héréditaires au gène "fonctionnel",
séquence de nucléotides, exprimé par la cellule
sous forme d'une chaîne polypetidique. Puis, par une
dérive dont on peut essayer de retrouver l'origine, les
gènes fonctionnels ont été assimilés aux
gènes héréditaires associés aux
caractères des organismes. Nous critiquerons cette
démarche en prônant un retour de la
prééminence de la biologie sur la chimie, du moins pour
ces questions sur le vivant.
Il est difficile d'arriver à savoir
quand a pris forme la théorie de l'information
génétique telle qu'elle est comprise par nombre de nos
contemporains. Il semble que certains physiciens et chimistes y
soient pour beaucoup (il est intéressant de noter que nombre
des chercheurs cités dans cette page ont commencé par
des études de physique et de chimie et ne sont venus à
la biologie moléculaire que plus tard). Max Delbrück
(1906-1981) qui fit partie du groupe du phage, et qui proposa
notamment une interprétation quantique des mutations qui
représenteraient un saut entre deux états stables du
gène, semble avoir influencé Erwin Schrödinger
(1887-1961) qui dans son livre "Qu'est-ce que la vie ?",
publié en 1944, parle déjà de programme
génétique (?) en ces termes "ces chromosomes (...)
qui contiennent sous la forme d'une espèce de code le
modèle intégral du développement futur de
l'individu et de son fonctionnement dans l'état adulte"
(cité dans "La
naissance de la biologie moléculaire").
On peut aussi souligner le rôle de
Linus
Pauling (1901-1994) qui, en plus de ses
découvertes fondamentales en chimie publiées notamment
dans "La nature de la liaison chimique" en 1939 et "Chimie
générale" en 1947, est celui qui a montré le
rôle essentiel des liaisons faibles en biochimie (une bonne
part de la compréhension moléculaire que l'on a du
vivant actuellement est due à ses idées), mais il est
aussi le découvreur des hélice alpha et feuillets
béta des protéines avec Corey en 1951, ou encore celui
qui proposé, en 1965, avec Emile Zuckerkandl, le concept
d'horloge moléculaire.
C'est Archibald
Garrod
(1857-?) qui exprime la première fois la possible relation
entre un gène et une enzyme. Il travaille sur une anomalie
métabolique humaine: l'alcaptonurie, qui affecte le
métabolisme de la tyrosine et de la phénylalanine
(voir
phénylcétonurie). Il
propose en 1909 dans un article intitulé "Les erreurs
innées du métabolisme", de justifier des
déficiences enzymatiques héréditaires de l'homme
(albinisme, cystinurie, pentosurie...) par des anomalies
génétiques dues à l'inactivation de gènes
codant pour certaines enzymes. Mais c'est
George Wells Beadle (1903-1989)
et
Edward Tatum qui, en 1941,
établissent de façon expérimentale cette
relation chez les mutants métaboliques de Neurospora
crassa.
Expérience (?) de Beadle et Tatum sur
Neurospora crassa
(datée de façon
erronée de 1941 dans...
Belin, Spécialité SVT,
2002, p 145)
Une page
complémentaire est
accessible afin de présenter avec davantage de
véracité historique et une réflexion
plus poussée ces résultats.
|
|
Beadle et Tatum isolent des souches mutantes
(que l'on qualifierait d'auxotrophes)
de N. crassa qui ne se développent pas sans
apport supplémentaire de tryptophane (un aa qu'elles
sont d'habitude capables de synthétiser) obtenues par
irradiation aux rayons X (ce que l'on
appellerait maintenant une mutagenèse
dirigée). Plusieurs souches sont obtenues pour
lesquelles l'apport de tryptophane peut être
remplacé par celui d'acide anthranilique ou encore
d'indole. Ces souches sont ensuite croisées entre
elles.
|
|
+ désigne une croissance
du mycélium sur chaque milieu
considéré
|
groupe 1
|
groupe 2
|
groupe 3
|
|
milieu minimum (MM)
(sucres, sels minéraux,
biotine (vitamine)...)
|
-
|
-
|
-
|
|
MM + acide anthranilique
|
+
|
-
|
-
|
|
MM + indole
|
+
|
+
|
-
|
|
MM + tryptophane
|
+
|
+
|
+
|
|
|
L'interprétation de Beadle et Tatum consiste dans
l'association d'un déficit métabolique
(transmissible héréditairement avec les lois
de l'hérédité chromosomique) avec une
mutation dans un gène qui devient alors une
unité fonctionnelle associé à la
synthèse d'une enzyme. La connaissance d'une
chaîne métabolique conduisant au tryptophane
date aussi de cette époque. Elle a cependant
été précisée car elle est
très complexe et ne présente pas une seule
voie, c'est un réseau
métabolique. La synthèse du
tryptophane est très bien connue par exemple chez les
bactéries et a fourni un exemple de régulation
génétique classique (opéron
trp).
|
Ces croisements sont bien sûr
facilités par le fait que les mycélium sont
haploïdes. La phase diploïde est réduite au
zygote, obtenu à la suite de la fécondation de
deux mycéliums compatibles, qui subit très
rapidement une méiose.
|
+ désigne une croissance sur MM
du mycélium issu de la spore
obtenue par croisement des mycéliums de
chaque groupe
|
groupe 1
|
groupe 2
|
groupe 3
|
|
groupe 1
|
-
|
+
|
+
|
|
groupe 2
|
+
|
-
|
+
|
|
groupe 3
|
+
|
+
|
-
|
Ces croisements sont ce que l'on apelle des tests de
complémentation. Si les mutations sont
situées dans la même unité fonctionnelle
(le même gène donc), la fécondation de
deux souches donne des mycéliums qui possèdent
la même déficience métabolique que les
souches croisées. Si les mutations sont situés
dans des gènes différents (mais qui conduisent
au même phénotype), les défisciences
peuvent se compenser (les étapes métaboliques
se complètent ou se complémentent) et l'on
peut obtenir un mycélium qui ne présente plus
de défiscience apparente.
|
|
Les trois groupes (1, 2 et 3) sont des souches qui sont
auxotrophes au Tryptophane. Mais chacune présente une
mutation dans un gène spécifique puisque
toutes complémentent. La chaîne
métabolique la plus simple que l'on peut proposer
pour interpréter ces résultats est
Précurseur ----(1)----> acide
anthranilique ----(2)----> indole ----(3)---->
tryptophane
Chaque gène (unité mutable) étant
associé à la fonction enzymatique
(unité fonctionnelle) permettant le passage d'une
substance à l'autre.
|
C'est Oswald
Avery (1877-1955) et
ses collègues Colin McLeod et McLyn McCarthy, qui
réussisent à purifier le facteur transformant du
pneumocoque après d'innombrables tentatives infructueuses de
nombreux chercheurs pendant 10 ans. Leur publication date de
1944.
|
Expérience d'Avery, McLeod et McCarthy,
1944
|
|
Ils réalisent un extrait chimique assez bien
purifié de SIII (voir expérience de Griffith)
contenant essentiellement des acides nucléiques (ARN
et ADN) et un peu de protéines. Ils ajoutent une
petite quantité de RII vivantes à une grande
quantité d'extrait de SIII. Ils obtiennent des SIII
vivantes. S'ils ajoutent à l'extrait de la trypsine
ou de la chymotrypsine (enzymes coupant de nombreuses
protéines) le résultat est le même.
S'ils ajoutent une ribonucléase à l'extrait le
résultat est identique. Par contre la transformation
n'a pas lieu s'ils ajoutent de la
désoxyribonucléase. C'est donc bien l'ADN
qui est le principe transformant.
|
En analysant ces deux
expériences dans une perspective moderne, on a voulu avoir la
preuve que l'ADN est une molécule transmissible et
suceptible d'être utilisée par une cellule
procaryote pour modifier une de ses caractéristiques
structurales : ici la synthèse d'une
capsule. Je pense
qu'à partir de la publication originale qui est accessible sur
internet on peut affirmer que cette interprétation est plus
que douteuse Voir une page
complémentaire
A partir de 1937, Max
Delbrück (1906-1981) travaille sur le bactériophage et
fonde ce qui va devenir la "Groupe du phage" avec l'arrivée en
1941 de Salvador Luria puis de Alfred Hershey en
1943. Une publication marquante de ces années est
l'expérience complémentaire de
Alfred
Hershey et
Martha
Chase de 1952 qui confirme le
rôle de l'ADN comme porteur de l'information
génétique du phage T2.
|
Expérience de Hershey et Chase,
1952
|
|
Le 32P est utilisé comme traceur pour l'ADN et le
35S pour les protéines. L'expérience utilise
des bactériophages T2 qui se reproduisent dans une
bactérie: Escherichia coli.
Deux lots de virions sont incubés en présence
d'E. coli: le lot 1 possède de l'ADN marqué au
32P et le lot 2 des protéines de la capside
marquées au 35S. Chaque culture est ensuite
mixée et centrifugée.
Pour le lot 1, c'est le culot qui est radioactif (30% de la
radioactivité initiale), alors que c'est le
surnageant du lot 2 qui est radioactif.
Les culots sont mis en culture et produisent des virus. On
observe une radioactivité de 30% de la
radioactivité initiale dans le lot 1
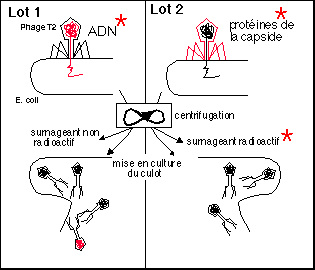
Lors de l'infection virale étudiée, c'est
donc bien l'ADN du phage T2 (et non les protéines de
la capsule) qui contient l'information
génétique et est transféré
à la cellule hôte. Cet ADN contient
l'information nécessaire à la
synthèse de nouveaux virions par la cellule
hôte.
|
Toujours dans la même
optique cette expérience prouve que l'ADN contenu dans les
virus et injecté dans les cellules hôtes
bactériennes peut être utilisé par celles-ci pour
produire de nouvelles particules virales.
Cette expérience semble avoir eu un
retentissement très grand notamment du fait qu'elle fut
publiée à peu près en même temps que le
modèle de la structure de la molécule d'ADN en 1953 par
James
Dewey Watson (1928-) et
Francis
Harry Compton Crick (1916-). Le
modèle en double hélice a été
élaboré à partir de la composition chimique de
l'ADN, notamment grâce aux travaux de Erwin Chargraff (1905-)
(qui avait montré que, pour toute molécule d'ADN, le
nombre de molécules d'adénine est égal au nombre
de molécules de thymine (A=T) et que celui de cytosine est
égal à celui de guanine (C=G)), des clichés de
diffraction X d'ADN cristallisé obtenus par
Maurice
Wilkins et de
Rosalind
Franklin décédée
prématuremment (1920-1958) (clichés publiés en
1953 dans le même numéro de nature que celui
présentant le modèle de Watson et Crick et
différenciant deux formes de la molécule: forme A et B
(hydratée)), et des clichés de microscopie
électronique qui montraient une molécule de 2 nm de
diamètre. Watson et Crick proposent déjà dans
leur article de 1953 un modèle d'appariement des bases pour la
réplication de la molécule.
Mais ce qui paraît maintenant fondamental
pour la direction qu'a prise la biologie moléculaire à
partir des années 1940 c'est surtout le rejet de toutes les
explications finalistes au sens de d'orientées
par la vie, phénomène propre du vivant et
irréductible à la chimie. Le hasard, qui n'est
pas sans rappeller la réversibilité des lois physiques,
devient la seule EXPLICATION ACCEPTÉE pour tout
DÉTERMINISME (ce qui, philosophiquement est un comble,
voir par exemple la
page sur
les niveaux d'organisation du vivant,
les biologistes ayant toujours préféré la
formulation utilisant les 3 grandes fonctions; nutrition, relation et
reproduction, que l'on peut qualifier d'autonomie ou de travail; en
ce qui concerne la finalité on peut lire le petit livre de
Dominique LETELLIER: La question du hasard dans l'évolution;
la philosophie à l'épreuve de la biologie, L'Harmattan,
Col. Ouverture philosophique, 2002, voir aussi le début
du
cours de TS). La domination des
physiciens et chimistes américains d'adoption et passés
au vivant grâce au soutien d'instituts privés comme le
Rockfeller Center n'y est sans doute pas étrangère.
Le premier article que l'on rattache à cette vision semble
être celui de Luria et Delbruck (Luria, S. E., and M.
Delbrück, 1943. Mutations of bacteria from virus sensitivity
to virus resistance. Genetics, 28: 491-511 - disponible
à l'adresse: http://www.esp.org/foundations/genetics/classical/holdings/l/slmd-43.pdf).
Il me semble qu'il faut séparer deux
démarches, mêmes si historiquement, elles ont
été mêlées :
- l'identification de l'ADN comme support de
l'information conduisant à la synthèse des
protéines et qualifiée d'information
génétique ; étant donné le but de
ces pages, cette partie ne sera pas reprise ici ; je renvoie par
exemple à Gènes, partie 2 : transformer les
gènes en protéines, pp 95-158 ; il est bien
évident que selon le point de vue adopté les
mécanismes moléculaires n'ont pas la même
signification. De même les données encore très
partielles sur la structuration de cette information : voir par
exemple les parties 3, 5 7 de Gènes.
On peut donc affirmer désormais qu'un gène
est une portion d'ADN de séquence spécifique qui
code pour la synthèse d'une molécule
fonctionnelle: la plupart du temps un polypetide mais parfois
aussi un ARN. Les gènes sont toujours
considérés comme organisés
linéairement sur les chromosomes mais ils sont
morcelés (maturation des ARNm, découverte des
introns et des exons chez quasiment tous les eucaryotes) et
peuvent se chevaucher. Les gènes possédent des
séquences de début et de fin dont l'étude est
loin d'être finie, ils peuvent se déplacer le long
des chromosomes (gènes "sauteurs"). Certains gènes
sont portés par de l'ADN non chromosomique (plasmides) ou
extranucléaire (mitochondries, chloroplastes).
On peut signaler la notion de cistron qui étant une
sous-unité fonctionnelle du gène
déterminée à partir des mutants: deux mutants
présentant des défisciences métaboliques
individuelles complémentaient, c'est-à-dire qu'un
individu avec un génotype issu de la réunion de leur
génome muté ne présentait plus de
défiscience. Actuellement, cette notion, inutilement
surajoutée au gène, semble être
abandonnée. Il suffit de présenter un cistron comme
un gène.
Les gènes sont des unités fonctionnelles et non plus
héréditaires. La composante
héréditaire du gène est maintenant inscrite
dans ce que nous savons de l'hérédité
chromosomique. Le gène n'est plus une unité
héréditaire attachée à un
caractère.
Il faut aussi souligner une confusion de vocabulaire sur le code
génétique. Une des caractéristiques de
l'ambiance moderne est justement de refuser un sens
véritable et immuable à un nom. C'est inadmissible
en science. Il faut être ferme là-dessus. Le code
génétique désigne la correspondance entre
les codons de l'ARN messager et les acides aminés des
protéines. Toute autre notion doit être
exprimée avec un autre nom. Notamment la correspondance
(qui n'est que supposition) entre une séquence d'ADN et une
étape du développement, ou l'apparition d'un
caractère...Il ne faut pas accepter que le sens soit
étendu au programme génétique.
- la recherche de la liaison entre
l'information génétique définie ci-dessus et
les caractères d'un organisme, c'est à dire la
signification même de cette information. Ce sont juste
quelques idées relatives à ces questions que je
souhaite développer ici et dans une optique bien
particulière exprimée dans les pages sur la
diversité du
vivant.
Les gènes
fonctionnels sont des séquences d'ADN manipulées par la
cellule et codant pour un produit fonctionnel (polypeptide,
ARN...)
Actuellement, la notion d'information
génétique est souvent confondue avec la notion,
tout à fait différente de programme
génétique. Si la première est tout à
fait justifiée au vu des résultats de la
génétique, la seconde est pour le moins contestable et
ne semble reposer sur aucun résultat probant (voir
cours de seconde, partie
II et les commentaires
du programme de seconde). Je me suis
aussi efforcé de présenter une interprétation
critique des résultats de la génétique du
développement vulgarisés (reposant tous sur la notion
de programme génétique de développement)
accessibles aux enseignants du secondaire dans un longue
page sur le
développement.
Dans le cadre de l'union entre théorie
héréditaire et théorie moléculaire, on
peut dire pour simplifier qu'un gène moléculaire
dominant (ce qui ne veut rien dire: il faudrait employer le terme
d'allèle associé à la forme d'un gène
moléculaire) est associé à un gain de fonction
et un gène récessif est associé à une
perte de fonction.
|
Résumé de l'étape 3
|
Garrod, Beadle et Tatum, Griffith, Avery, Mc Leod et
Mc Carthy, Hershey et Chase, Watson et Crick, Wilkins et
Franklin, Schrödinger et Pauling
|
Le gène est une unité fonctionnelle :
c'est une portion d'ADN de séquence spécifique
qui code pour la synthèse d'une molécule
fonctionnelle. La mutation est un changement de
séquence de l'ADN.
Certains biologistes moléculaires superposent
la notion de gène, unité fonctionnelle et
celle de gène, particule héréditaire.
C'est cette notion qui est la plupart du temps
enseignée dans le secondaire. L'ADN, scindé en
sous-unités fonctionnelles, les gènes, est
la molécule de
l'hérédité. Les lois de transmission de
l'ADN sont les lois de l'hérédité. On
passe donc à un modèle moléculaire
de l'hérédité. Certains vont
jusqu'à réduire l'individualité d'un
être vivant à l'originalité de son
génome.
Ces théories ont été
élaborées d'une part à partir des virus
et d'autre part à partir des procaryotes. Dès
que l'on s'adresse à des eucaryotes on revient au
modèle chromosomique qui n'est pas réductible
au modèle moléculaire.
|
Une théorie de
l'hérédité...
1. de l'information génétique
à l'information cytoplasmique et retour
Parler d'une information cytoplasmique qui
puisse être transmise de façon héréditaire
toute comme l'information génétique de l'ADN repose sur
des données expérimentales acquises dans les
expériences d'embryologie
(voir page
sur le développement).
Une autre voie, vraiment novatrice, a été ouverte
grâce à la connaissance des ARN, qui peuvent être
considérés non pas comme des produits de
l'activité des gènes et donc de l'ADN, mais comme des
molécules informatives, capables notamment de transmettre une
information à d'autres cellules et même de
s'intégrer à l'information génétique
d'une cellule procaryote. On sait que certains ARN ont des
propriétés catalytiques (ribozymes). D'autres ont donc
aussi la propriété de pouvoir manipuler l'ADN. La
cellule pouvant même transférer cette information de son
support d'ARN à un support d'ADN et l'intégrer ainsi
à son génome.
Voici des expériences peu médiatisées au sujet
de l'information portée par les ARN (pour les
références d'autres articles originaux voir
Beljanski, un novateur en biomédecine; concepts,
théories, applications; C.-G. NORDAU et M.S. BELJANSKI,
éd. EVI Liberty Corp, 2001):
Expérience(s) de M. Beljanski, M.S.
Beljanski, P. Manigaut, P. Bougarel (1971, 1972...1986)
ARN transformants
M. Beljanski, M.S. Beljanski, P.
Bougarel, ARN transformants porteurs de caractères
héréditaires chez Escherichia coli
showdomycino-résistant, C.R. Acad. Sci., 1971,
272, pp 2107-2110 (série D)
|
Schémas réalisés
d'après une description des expériences (in
Nordau et Beljanski) sans avoir eu accès aux
publications originales...
La Showdomycine, antibiotique extrait d'un champignon
(Streptomyces) d'extrême Orient est un
nucléoside naturel proche de l'uridine qui semble
donc pouvoir interagir avec les acides
ribonucléiques. E. coli développe
très facilement une résistance à cet
antibiotique.
Les ARN libérés dans le milieu par les
souches résistantes sont présents dans les
souches sauvages mais ils ne sont libérés que
chez les souches mutées. Les auteurs pensent que ces
ARN sont fixés normalement à l'ADN
(épisomes à ARN) et stabilisent ainsi
sa structure (M. Beljanski, M.S. Beljanski
et P. Bougarel, "Épisomes à ARN"
porté par l'ADN d' Escherichia coli sauvage et
showdomycino-résistant, C.R. Acad. Sci., 1971,
272, pp 2736-2739 (série D))
Dans la culture (4) il n'y a pas de transformation si
l'on ajoute une ribonucléase (enzyme dégradant
les ARN).
La croissance des souches mutées résistantes
est toujours légèrement supérieure
à celle des bactéries sauvages. Ceci est
interprété par à un relâchement
de la chaîne d'ADN qui favorise les synthèses
dans la souche mutée résistante.
Les ARN transformants ne s'hybrident que très peu
à l'ADN des souches sauvage ou mutante, il est donc
peu probable qu'ils proviennent de la transcription de
gènes. Leur origine pourrait être la
PNPase (PolyNucléotide Phosphorylase),
extraite des mutants (et des souches sauvages) et capable de
synthétiser des ARN synthétiques (poly AGUC)
dont les teneurs respectives en A,U, G et C ne
dépendent pas que de la teneur en chacune des bases
dans le milieu (si les 4 bases sont en quantité
égale, l'ARN synthétisé contient 2 fois
plus de bases puriques que pyrimidiques, comme dans le cas
des ARN transformants). Mais la PNPase peut aussi
synthétiser de l'ARN à partir d'un brin
matrice. In vivo la PNPase est toujours liée
à de petites quantitées d'ARN. Chez les
mutants Showdomycine résistants, elle est liée
à l'ARN transformant, ce qui suggère qu'elle
en est bien à l'origine, tout en l'utilisant comme
matrice.
La Showdomycine est inefficace sur l'activité de
l'ARN polymérase ADN dépendante.
Interprétations proposées...
|
|
Des souches d'Agrobacterium tumefaciens , une
bactérie bien connue pour sa capacité à
développer des cals (tumeurs
végétales), sont ensuite exposés in
vitro aux ARN transformants d'E. coli. Elles sont
alors transformées et perdent leur pouvoir
tumorigène tout en acquérant d'autres
propriétés (M. Beljanski, M.S.
Beljanski, P. Manigaut, P. Bougarel, Transformation of
Agrobacterium tumefaciens into a non-oncogenic species by an
Escherichia coli ARN, Proc. Nat. Acad. Sci. (USA), 1972,
69, pp 191-195). Cette transformation est
réellement l'acquisition d'une nouvelle information
génétique car ces bactéries
transformées ne gardent pas la trace de l'ARN
transformant. L'équipe va donc alors rechercher
comment une information venant de l'ARN a pu
s'intégrer au génome. Et c'est la
découverte de transcriptase réverse
(retrotranscriptase ou transcriptase inverse permettant la
synthèse d'ADN à partir d'une matrice d'ARN)
non seulement chez Escherichia coli
(M. Beljanski,
Synthèse in vitro de l'ADN par une transcriptase
d' Escherichia coli, C.R. Acad. Sci., 1972, 274, pp
2801-2804 (série D) et M. Beljanski, M.S. Beljanski,
RNA-bound Reverse Transcriptase in Escherichia coli and
in vitro synthesis of a complementary DNA, Biochemical
genetics, 1974, 12, pp 163-180))
mais ensuite chez Agrobacterium tumefaciens
(M. Beljanski, P. Manigault, Genetic
transformation of bacteria by RNA and loss of oncogenic
power properties of Agrobacterium tumefaciens. Transforming
RNA as template for DNA synthesis, Sixth Miles
International Symposium on Molecular Biology,. Ed. F. Beers
and R.C. Tilghman, The Johns Hopkins University Press,
Baltimore, 1972, pp 81-97; M. Beljanski,
Séparation de la transcriptase inverse de l'ADN
polymérase ADN dépendante. Analyse de l'ADN
synthétisé sur le modèle de l'ARN
transformant, C.R. Acad. Sci. , 1973, 276, pp 1625-1628
(série D)) puis chez Neurospora crassa
(S.K. Dutta, M. Beljanski, P. Bougarel,
Endogenous RNA-bound RNA dependant DNA polymerase
activity in Neurospora crassa, Exp. Mycology, 1977, 1,
pp 173-182) puis dans des
œufs de poisson (M. Beljanski, L.C.
Niu, M.S. Beljanski, S. Yan, M.C. Niu, Iron stimulated
RNA-dependant DNA polymerase activity from goldfish
eggs, Cellular and Molecular Biology, 34, 1988, pp
17-25), toujours par des équipes travaillant
avec M. Beljanski. La découverte d'une transcriptase
inverse chez un virus revient à Temin (1970).
L'ARN, avec les travaux de Beljanski, est reconnu
comme vraiment transformant c'est-à-dire
capable de transmettre une information
génétique au sens large ou
héréditaire chez les Procaryotes entre
bactéries de la même espèce
(transformer une souche bactérienne non
résistante en une souche résistante, de
façon stable) puis entre bactéries
d'espèces différentes (à partir
d'un ARN transformant d'E. coli, Agrobacterium
tumefaciens perd son pouvoir tumorigène,
grâce à l'insertion d'un nouvel ADN issu de la
transcription inverse de l'ARN transformant). L'information
génétique ou plutôt
héréditaire peut donc passer par l'ARN. Dans
le cas des épisomes à ARN, cette information
peut donc venir de l'ARN, être transmise sous forme
d'ARN, puis être intégrée au
génome sous forme d'ADN.
|
L'information génétique
peut donc aller de l'ADN à l'ARN mais aussi de l'ARN
à l'ADN de façon stable et transmettre ainsi une
propriété nouvelle intégrée au
génome.
Depuis d'innombrables travaux ont mis en évidence l'importance
des ARN chez les Procaryotes et les Eucaryotes, en association avec
l'ADN sous forme d'hybrides d'ADN-ARN (probablement issu d'une
"transcription inverse") , ou sous forme d'ADN d'origine non
nucléaire.
Le devenir des ARN transformants:
hybride ARN-ADN stabilisé, ARN libre actif,
intégré à l'ADN sous forme d'ADN, associé
à l'ADN sous forme d'épisome...
Ces travaux sont encore très partiels et
presque limités à des bactéries. Mais on peut
sans aucun doute continuer à travailler dans ce sens (ceci est
un appel à des vocations de jeunes chercheurs). Il existe de
nombreux cas où les cellules synthétisent de grandes
quantité d'ARN, tout particulièrement dans les
ovocytes, surtout chez les amphibiens. Ces ARN ont naturellement
été considérés comme des ARN messagers.
Mais les travaux de Beljanski pourraient nous faire voir tout
différemment ces synthèses (voir page
sur le développement): des ARN
courts pourraient ainsi constituer des épisomes ou des
matrices pour des fragments d'ADN qui s'inséreraient dans le
génome puis qui seraient ainsi transmis dans certaines
lignées.
2. l'information génétique
manipulée par le cytoplasme
M. Beljanski a ensuite surtout porté ses
recherches dans le domaine médical dans un but
appliqué: trouver des molécules qui stabilisent l'ADN
dont la déstabilisation (rupture des liaisons faibles et
ouverture de la double hélice) semble bien être la cause
des cancers (voir page
sur le cancer). Ses travaux l'ont ainsi
conduit à isoler des ARN-fragments, c'est-à-dire
de petits ARN courts (quelques dizaines de nucléotides) issus
de la dégradation d'ARN ribosomiaux notamment (ARN
amorceurs puis ARN antisens) qui lui ont permis de
stabiliser l'ADN de cellules cancéreuses de mammifères
et puis de cellules cancéreuses humaines. Je renvoie à
l'ouvrage Beljanski, un
novateur en biomédecine; concepts, théories,
applications; C.-G. NORDAU et M.S. BELJANSKI, éd. EVI Liberty
Corp, 2001 pour une bibliographie.
Cependant, M. Beljanski a tout de même eu le temps de
développer une théorie passionnante sur l'expression de
l'information génétique qui a été reprise
par R. Chandebois dans son ouvrage Le gène et la forme
et dans une moindre mesure dans Comment les cellules
construisent l'animal (voir bibliographie). En voici un bref
aperçu:
Théorie de la
régulation de l'expression de l'information
génétique ADN dépendante par
stabilisation-déstabilisation de l'ADN
(M. Beljanski, Activation et
inactivation des gènes. Incidence en
cancérologie., Aspect de la recherche,
Université de Paris Sud, 1985, pp 56-62; M.
Beljanski, The regulation of DNA Replication and
Transcription. The Role of Trigger Molecules in Normal and
Malignant gene expression, Experimental Biology and
Medecine, vol. 8, Karger, 1983, pp 1-190)
Théorie reprise et élargie au
développement par Rosine Chandebois dans Le
gène et la forme (ou la démythification de
l'ADN), Rosine Chandebois, 1989, Ed. Espaces 34, p 67
à 74.
|
Des substances chimiques comme les
cancérogènes (en violet)
peuvent stabiliser ou déstabiliser
l'hélice d'ADN, ce qui rend plus accessible ou au
contraire moins accessible certains gènes aux
complexes enzymatiques de réplication (ADN
polymérase) et de transcription (ARN
polymérase). Ce schéma est juste à but
d'illustration et ne correspond à aucun
résultat expérimental précis
(d'après Beljanski, un novateur en
biomédecine, p 42).
Plus la transcription est importante et rapide plus les
chaînes d'ADN s'écartent au site de
transcription mais se ressèrent sur les sites
adjacents, qui deviennent quiescents , par force (c'est un
phénomène mécanique), par manque
d'espace pour l'accès de l'ARN polymérase aux
gènes. Les histones pourraient vérouiller les
sites non accessibles aux enzymes de transcription où
limiter leur transcription en-dessous d'un certain seuil
(métabolisme larvé, résiduel ou encore
"de luxe").
Ainsi des protéines, des ARN, des substances
très variées, comme les
cancérogènes, pourraient venir manipuler
spatialement (mécaniquement) l'ADN et
contrôler ainsi l'expression de ses
gènes, non pas par des produits isssus de
l'activité de tel ou tel gène
"régulateur", mais de façon beaucoup plus
complexe (car moins localisé et plus
interpénétré) mais aussi beaucoup plus
simple (en échappant aussi au concept du gène
régulateur, réglé lui-même par un
autre gène et ainsi de suite), par des produits
très variés du métabolisme ou de la
communication entre cellules.
Cette théorie est un grand soutien à
l'idée d'information génétique,
manipulée par une information cytoplasmique,
elle-même sous la dépendance d'une information
extracellulaire.
Mais il est sûr qu'il y a encore beaucoup de
travail à faire. Trouver des molécules
régulatrices synthétisées sans le
concours de gènes spécifiques comme pour les
cancérogènes, est certainement une voie de
recherche ouverte. Cette théorie n'en est encore
qu'à ses balbutiements.
|
3. vers une théorie de
l'hérédité cellulaire
Quelques points non ordonnés pour
l'instant :
- D'abord il y a un problème de
signification :
Ce n'est pas le même type de transmission
héréditaire que l'on étudie quand on
s'intéresse à la pénétration de l'ADN
d'un bactériophage dans une bactérie, ou un
échange d'ADN par pont cytoplasmique entre deux
bactéries, ou encore une mitose ou une méïose
chez un eucaryote (et encore faut-il accepter de composer avec les
caractéres individuels de chaque reproduction dans chaque
espèce...). Il y a certes transmission de matériel
génétique mais celui-ci n'a pas forcément la
même signification dans chaque cas. Je renvoie par exemple
à l'article : Echanges de gènes entre
bactéries, Robert MILLER, 1998, La Recherche, 245, mars
1998, 60-65. Je propose de ne pas considérer les
échanges de gènes de ce type comme faisant partie
des fonctions de reproduction mais bien des fonctions de relation.
Echanger de l'ADN est, pour ces bactéries, un moyen de
communiquer. D'ailleurs les lois de recombinaison des gènes
bactériens n'obéissent pas du tout à
l'hérédité mendélienne. De la
même façon, les virus n'étant pas des
êtres vivants (voir discussion dans les pages sur la
diversité du
vivant), les échanges d'ADN
par les virus font partie des relations entre organismes
(cellulaires donc).
- Ensuite, il y a un problème
d'échelle :
La linéarité des gènes
héréditaires (mise en évidence par
l'hérédité morganienne) n'est pas du
même ordre de grandeur que celle des gènes
fonctionnels mis en évidence par la biologie
moléculaire. Quand on change d'ordre de grandeur il est
fréquent que l'on change de mécanisme ou
plutôt que l'on soit obligé de changer de
modèle pour expliquer les mécanismes du
réél.
Je remarque que Lewin fait un raisonnement inverse dans
Gènes p 73 : "Nous n'avons aucune raison de croire que
la situation serait différente chez les eucaryotes si nous
étions à même de réaliser une
cartographie génétique fine".
Le gène, unité d'information génétique
est considéré comme stable, voire immuable,
lorsqu'on l'utilise dans une empreinte
génétique ou une cartographie de
restriction. Alors que l'on sait bien que le génome est
parfois mobile et que de nombreuses modifications du génome
n'altérent pas le fonctionnement cellulaire.Comment se fier
à la position de sites de restriction pour
caractériser l'ADN d'un individu alors que l'on ne sait
même pas si toutes les cellules possédent bien
effectivement les mêmes séquences (on ne l'a pas
testé puisque le génome d'une seule cellule humaine
n'est même pas séquencé) : on part du postulat
de départ que l'information génétique est
répliquée sans aucune erreur et entiérement
à chaque division depuis la cellule oeuf. Ce qui est
peut-être vrai pour une bactérie ne l'est pas
forcément pour l'homme.
La notion d'unité de recombinaison n'a pas du tout le
même sens si l'on s'adresse aux procaryotes et aux
eucaryotes. La recombinaison proposée comme
interprétation de la liaison partielle entre deux
caractères n'est pas du forcément la même que
la recombinaison dite généralisée
étudiée expérimentalement chez quelques
procaryotes et quelques eucaryotes. La manipulation du
génome, la plupart du temps in vitro, ne justifie pas
l'interprétation de la liaison partielle.
Tableau essayant de rendre compte des
différentes échelles d'étude du
"génome"
|
"organisme"
|
taille du "génome" en (paires
de) bases
|
matériel "génétique"
|
nombre de gènes
fonctionnels estimé par la biologie
moléculaire
|
nombre d'unités de carte
factorielle connues
(carte génétique ou
statistique en (paires de) bases
|
taille théorique de
l'unité de recombinaison
( = taille du génome / nombre de recombinaisons
connues) en (paires de) bases
|
distance moyenne théorique
entre les sites de recombinaison
(= taille du génome / taille de l'unité de
recombinaison) en (paires de) bases
|
|
virus
|
virus de la grippe
|
13 500
|
ARN simple brin
|
12
|
|
|
|
|
bactériophage T4
|
165 000
|
ADN double brin
|
~ 200
|
800
|
200
|
10 000
|
|
procaryotes
|
mycoplasme
|
< 1 000 000
|
750 ?
|
|
|
|
|
Escherichia coli
|
4 200 000
|
2 000 ?
|
1750
|
2400
|
120 000
|
|
eucaryotes
|
Saccharomyces cerevisiae (protiste :
levure)
|
20 000 000
|
4 000 ?
|
4200
|
5000
|
250 000
|
|
Nématode
|
800 000 000
|
5 000 ?
|
320
|
250 000
|
12 000 000
|
|
Drosophila melanogaster
(Invertébré, Arthropode)
|
140 000 000
|
10 000 ?
|
280
|
500 000
|
25 000 000
|
|
Souris
|
3 000 000 000
|
25 000 ?
|
1700
|
1 800 000
|
90 000 000
|
|
Homme
|
3 300 000 000
|
50 000 ?
|
3300 ??
|
1 000 000 ??
|
50 000 000 ??
|
|
Quelques chiffres...
- 120 000 paires de bases = taille d'une bande
colorée sur un chromosome humain (estimée
à partir d'une coloration à haute
résolution de 550 bandes pour un chromosome moyen
de 65 000 000 paires de bases = 46 chromosomes pour 3,3
milliards de paires de bases)
- 25 000 000 de paires de bases = taille estimée
de la plus petite distance entre deux sites qui ne
recombinent pas statistiquement par crossing-over chez la
drosophile (la méthode est inapplicable à
l'homme)
- un fragment de 50 paires de bases n'est quasiment pas
récupérable dans un gel d'agarose
(Gènes, p 76), on peut estimer à quelques
centaines de milliers de paires de bases les plus petits
fragments de restriction manipulables (je ne suis pas
sûr de ce chiffre mais je n'ai trouvé aucune
donnée précise à ce sujet). A ce
titre l'exercice du livre de Terminale S Bordas
spécialité n°7 p34 probablement repris
de Gènes p 75 n'est pas une approche
réelle, c'est un exercice THEORIQUE ce qui est
franchement déconcertant ; les chiffres sont
donnés en paires de bases alors qu'il devraient
l'être en centaines de milliers de paires de
bases.... par contre, avec les méthodes de
séquençage d'ADN par
électrophorèse sur gel d'acrylamide, on
arrive à faire migrer des fragments de quelques
centaines de paires de bases ... (Gènes p85)
|
|
Vers une nouvelle étape
|
Une théorie complète de
l'hérédité doit tenir compte des trois
niveaux d'information du vivant : le niveau
génétique, le niveau cytoplasmique (qui
manipule l'information génétique) et le niveau
extracellulaire (l'environnement).
Par exemple le zygote humain posséde bien les
trois types d'information : l'information
génétique qui résulte de la
réunion des informations génétiques des
deux gamètes haploïdes (un exemple
d'application étant une anomalie chromosomique qui
provoque des anomalies graves sur le développement) ;
l'information cytoplasmique qui résulte de
la fusion inégale des cytoplasmes de l'ovocyte II et
très partiellement (voir pas du tout) du
spermatozoïde (un exemple la mettant en
évidence peut par exemple être fourni par
l'étude des cas de certains vrais jumeaux qui, par
division d'un massif cellulaire correspondant à un
individu, donnent deux individus, identiques par certains
traits morphologiques mais pas au niveau de l'individu
biologique bien évidemment, ce qui reviendrait
à dire qu'ils ne sont qu'une seule personne...);
l'information extra-cellulaire qui résulte des
interactions entre les gamètes et les cellules
présentes lors de leur maturation, puis de l'oeuf et
des cellules voisines du lieu de la fécondation puis
du lieu de développement de l'embryon
(information clairement démontrée par les
expériences d'embryologie expérimentale...
pour donner un exemple classique tiré du livre de
Rosine Chandebois : le gène et
la forme (p 62): lorsqu'on fournit à un
épithélium oesophagien de poulet de la
vitamine A, il cesse de sécréter de la
kératine et sécréte alors du mucus ; ou
pour rester dans le domaine de la biologie humaine, si une
fécondation a lieu en un autre endroit que la partie
terminale des trompes, le développement ne se
poursuit pas...).
|
En guise de conclusion
provisoire, juste quelques réflexions, à
améliorer, je n'en doute pas.
Le premier point essentiel concerne l'organisme
: la génétique des procaryotes ne peut pas
être assimilée à celle des eucaryotes, c'est
un problème d'ordre de grandeur essentiel (voir tableau plus
haut).
Si l'on considère par exemple chez la drosophile deux
gènes classiquement considérés comme liés
(gènes b pour black (corps noir) et vg pour (ailes)
vestigiales) portés par le chromosome n°2. Ces
allèles sont définis en terme d'unités
mutables mais certainement pas en terme d'unité codant
pour un produit fonctionnel, même si évidemment, on
connaît des protéines codées par cette immense
partie du chromosome polyténique drosophilien...De la
même manière l'allèle sauvage est simplement
défini comme le gène non muté mais certainement
pas comme une unité fonctionnelle. Que signifie dans ce cas,
pour le chromosome et le gène drosophilien une liaison
chromosomique ? Il semblerait que l'on puisse répondre que
deux unités mutables liées correspondent à deux
caractères qui apparaissent toujours simultanément
chez l'adulte et qu'ils peuvent être reliés
matériellement à deux zones proches d'un chromosome
donné. Il y a donc un lien, exprimé au cours de
l'embryogenèse, entre ces deux caractères, ce lien
passant par l'expression de produits gouvernés par les
gènes en question (sans que l'on ait pour l'instant une
idée précise de la relation entre le génotype et
le phénotype ... de l'adulte... voir à ce propos la
page du nouveau
cours de 1ère S). Ce lien est
donc toujours métabolique car il s'exprime à la fois au
niveau cytologique et au niveau extracellulaire puisqu'on le retrouve
au niveau de l'organisme adulte.
De la même manière que signifie alors un crossing-over ?
Les deux caractères portés par chacun des parents
séparément avec deux allèles différents
se retrouvent associés (sont exprimés ensemble) chez un
descendants. Reprenons l'exemple ci-dessus. Une drosophile
mutée à ailes vestigiales et corps noir de souche
"pure", c'est à dire qui se reproduit toujours en donnant des
descendants qui présentent les mêmes mutations
présente donc une modification probablement matérielle,
transmise héréditairement, et qui repose de
façon certaine, au moins sur deux zones bien
spécifiques du chromosome n°2. Si l'on regarde maintenant
l'organisme entier et que l'on ne s'intéresse plus uniquement
à ses chromosomes, on a donc des mouches à corps
noir et ailes vestigiales qui se reproduisent entre elles et donnant
toujours des mouches à corps noir et ailes
vestigiales....
Que doit-on garder de Mendel ? Si
l'hérédité mendélienne doit probablement
être considérée comme l'exception il n'est reste
pas moins que la liaison chromosomique entre les caractères
et leur support matériel -le gène matériel-
semble être une base incontournable. Mais bien
évidemment l'expression du gène -le gène
fonctionnel-, connaissance acquise grâce à la biologie
moléculaire, n'est pas du tout univoque. La liaison entre un
gène et un caractère peut nécessiter
l'expression d'un autre gène, lié physiquement ou non
(on pourrait dire qu'il est lié métaboliquement).
Que doit-on garder de Morgan ? Je me méfie de plus en plus du
modèle "drosophilien" mais la disposition linéaire
des gènes semble être aussi acquise. Par contre
le crossing-over pourrait être écarté non pas
comme un mécanisme qui n'existe pas (c'était le jeu
! : voir cours de TS sur un jeu
sans crossing-over) mais bien comme
un mécanisme, parmi d'autres mécanismes, de
recombinaison site-spécifique , celui-ci n'étant
probablement pas un mécanisme essentiel à la
transmission des gènes. De la même manière, et de
façon certes très audacieuse (!!!) on pourrait aussi
écarter la méiose telle que l'on la conçoit ;
une succession inamovible de deux divisions
précédée par une longue étape de
maturation pendant laquelle on lieu les crossing-over. Une nouvelle
lecture consisterait à dire que la période de
maturation sexuelle est la seule étape vraiment originale et
spécifique (avec la fécondation ou le
déclenchement de la division de l'œuf) de la reproduction
sexuée. Il n'existerait alors qu'un seul type de division
cellulaire qui serait la mitose. (Voir
cours de TS
sur la reproduction et page
sur la
méïose).

retour accueil
/ tableau histoire
des sciences