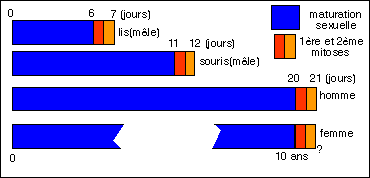
Comparaison entre les temps de maturation de la cellule sexuelle (gonie) et la durée des deux divisions
(appelées mitoses ici: mitose sexuelle et mitose ganètique) qui lui succèdent chez quelques organismes.
retour accueil, page sur la génétique
Cette page est une proposition ouverte (même si je procède par affirmations) de reconsidérer la méïose comme succession de deux divisions celullaires particulières et y substituer la notion de maturation sexuelle, qui précéderait dans le temps certaines des divisions cellulaires eucaryotes ("mitoses sexuelles et gamètiques") qui touchent les cellules de la lignée germinale.
Si l'on part de considérations cytologiques et si l'on se
réfère aux mécanisme connus sans
s'intéresser à la structure des chromosomes, il semble
bien n'y avoir qu'un seul type de division cellulaire. On pourrait
dire qu'il n'y a qu'une seule division cellulaire : la
mitose.
Si l'on part de considérations embryologiques les
modalités de divisions , l'orientation des fuseaux, la taille
des cellules filles par rapport à la cellule mère, la
répartition des organites dans les cellules filles....les
modalités de division cellulaire sont très
nombreuses chez les eucaryotes. La mitose n'est qu'un nom qui
désigne la division cellulaire, elle n'a pas la même
valeur, les mêmes conséquences selon les cellules qui la
subissent. Une mitose de renouvellement de cellules
épidermiques, une mitose de croissance dans l'os, une mitose
de segmentation embryonnaire, la première mitose de l'embryon
au stade une cellule...sans compter les mitoses incomplètes,
celles qui ne conduisent pas à la séparation des
cellules filles (c'est le cas le plus préquent dans les
divisions goniales), celles qui ne conduisent pas à la
division du noyau (c'est la cas de la maturation des
spermatozoïdes à partir des spermatides...ou de la
fragmentation des plaquettes à partir d'un
mégacaryoblase). Pourquoi ne pas considérer que les
cellules de la lignée germinale, constituent , dans cette
optique, une originalité.
Dans le cadre d'une théorie du
vivant non pas fondée sur l'information
génétique mais sur le comportement social
élémentaire de chaque cellule, la division
cellulaire, ou mitose est un des principaux processus qui
reflète l'autonomie du vivant. Lorsque l'autonomie de la
cellule n'est plus considérée de façon
cachée comme résultant de substances chimiques qui
actionnent des gènes, mais comme une véritable
composante de la cellule, au même type que sa
matière, on ouvre la porte à une interprétation
vraiment globale du mécanisme de la division cellulaire,
AUTONOME.
Selon les conceptions généralement en vigueur, les
étapes de la vie d'une cellule ont été
replacées dans un cycle, tout en sachant que certaines
étapes peuvent être très longues, voir
définitives, la cellule restant bloquée dans cet
état intermédiaire. On a donc cherché (et
trouvé) des déterminants pour les différentes
étapes du cycle cellulaire. Mais si l'on prend comme
donnée l'autonomie de division, les modalités (types
cellulaires impliqués, localisation, contraintes de forme ou
mécaniques...) deviennent des éléments
essentiels d'une théorie de la division cellulaire et les
substances chimiques deviennent à leur tour, non pas la
condition de l'autonomie cellulaire mais bien l'expression de
cette autonomie.
Que se passe-t-il donc de différent dans les cellules sexuelles ? D'une part il y a une période de maturation sexuelle qui est plus ou moins longue mais qui précède la mitose sexuelle. Classiquement on place ces phases dans la prophase de la première division de méiose. On distingue 5 stades inégaux ou non selon l'aspect des chromosomes qui sont alors visibles (leptotène, zygotène, pachytène, diplotène et diacinèse). Cette période de maturation est toujours longue par rapport aux divisions qui lui font suite (plus de 11 jours sur 12 en comptant les deux divisions aboutissant à la formation des spermatozoïdes chez la souris, 6 jours sur 7 pour la formation des grains de pollen chez le lis, plus de 10 ans si l'on considère l'ovocyte féminin, et plus de 20 jours sur 21 pour les spermatozoïdes humains).
C'est pendant cette longue période que l'on place
l'hypothétique crossing-over dont nous allons reparler.
C'est pendant cette longue période qu'a aussi lieu la
récupération de la mémoire cytoplasmique de
l'espèce dans le cadre de la théorie
de l'évolution de Mme Chandebois.
Si l'on se réfère à ce qui se passe lors de la division qui suit cette période de maturation sexuelle, il y a toujours séparation des chromosomes homologues et non des chromatides... pourquoi ne pas alors invoquer les fameux complexes synaptonémaux et les chiasma qui en résultent (?) pour justifier que dans la mitose qui lui fait suite, les chromatides de chaque chromosome restent aisément accolées alors que les chromosomes homologues sont tirés chacun d'un côté de la cellule ou de l'autre. La mitose qui fait suite à une maturation sexuelle, que l'on peut qualifier de mitose sexuelle sépare donc les homologues et non les chromatides de chaque chromosome. Si les cellules filles en restent là et ne se divisent plus on a des cellules à n chromosomes mais avec une quantité d'ADN identique à celle de la cellule de départ (classiquement notée Q).
Selon la place de cette maturation sexuelle et donc de la mitose sexuelle, on peut avoir une pré ou une postréduction (voir ci-dessous).
La division suivante peut se faire directement sans passer par une interphase avec notamment une phase S de synthèse, étant donné que les chromosomes de la cellule mère ont déjà deux chromatides qui se sont séparées (elles ne forment plus de complexe synaptonémal). La mitose qui suit, que l'on peut donc qualifier de mitose gamétique, sépare les chromatides de chaque chromosome homologue dans chaque cellule fille de la division précédente. On obtiendait ainsi des cellules haploïdes avec une quantité d'ADN de Q/2.
Conclusion
Ces lignes ne sont pas une recherche d'originalité mais sont
la conséquence du travail mental que l'on fait lorsque l'on
s'efforce de changer de paradigme. IL ne s'agit pas d'un savoir
scientifique, expérimental, sociologiquement reconnu, mais de
pistes qui pourraient à mon avis être suivies par la
jeune génération.
Dans ce sens, c'est une évidence de dire qu'une des
clés de la division cellulaire est sans aucun doute dans la
formation et la duplication des centrosomes. On sait provoquer chez
l'oursin des bourgeonnements de la cellule qui contiennent des
centrosomes mais pas de noyau (Biologie moléculaire de la
cellule, 3ème éd., p 914). Une recherche de
déterminismes de la division cellulaire, qui se ferait dans
l'optique de la nouvelle théorie de Mme Chandebois partirait
dans une toute autre direction que celle qui suit le paradigme
dominant (encore) actuellement. L'hérédité
stéréospécifique liée aux centrioles est
connue depuis 1960 (Biologie moléculaire de la cellule,
Alberts et al., 1995, Médecine-Sciences Flammarion, ch 16, p
820)...
Voici un exemple d'exercice de génétique réinterprété dans cette optique... je ne sais pas bien sûr si on peut faire de même dans beaucoup d'autres cas...
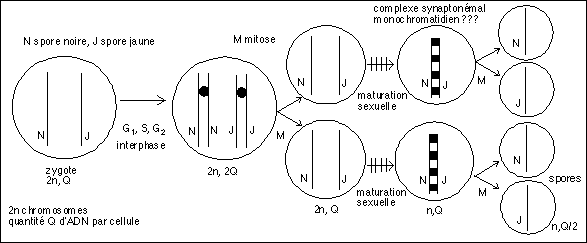
Un exemple pratique, revenons aux champignons haploïdes (Sordaria et autres Neurospora) du cours de génétique... on avait émis encore une autre hypothèse anticonformiste dans un jeu proposant la non existence du crossing-over. Bizarrement, on retrouve ici cette même idée. Pour expliquer les asques postréduites point n'est besoin du crossing-over. Il suffit donc de faire une subir à la spore zygotique une mitose (on passe de 2n chromosomes et une quantité SQ d'ADN dans l'unique cellule à deux cellules à 2n chromosomes et à une quantité Q d'ADN), puis une maturation sexuelle pendant laquelle il y a accolement des chromosomes homologues, alors monochromatidiens, puis une deuxième mitose (mitose sexuelle) qui sépare alors les chromosomes homologues dans chacune des cellules filles qui ne sont plus qu'à n chromosomes et avec une quantité d'ADN Q/2, comme dans une méiose classique. Dans ce cas le fait que la maturation sexuelle intervienne avant ou après la première mitose justifie les proportions d'asques pré- et postréduites dont la signification n'est plus la distance au centromère mais un tout autre phénomène cytologique.
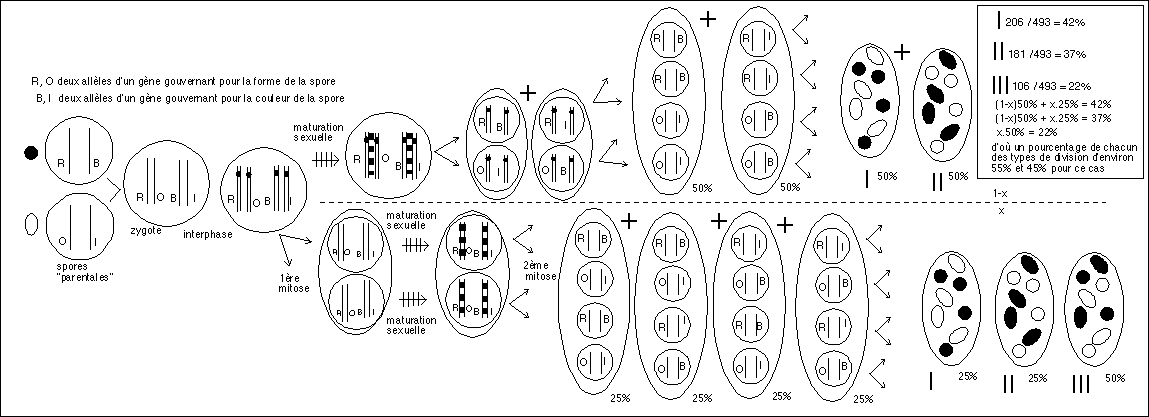
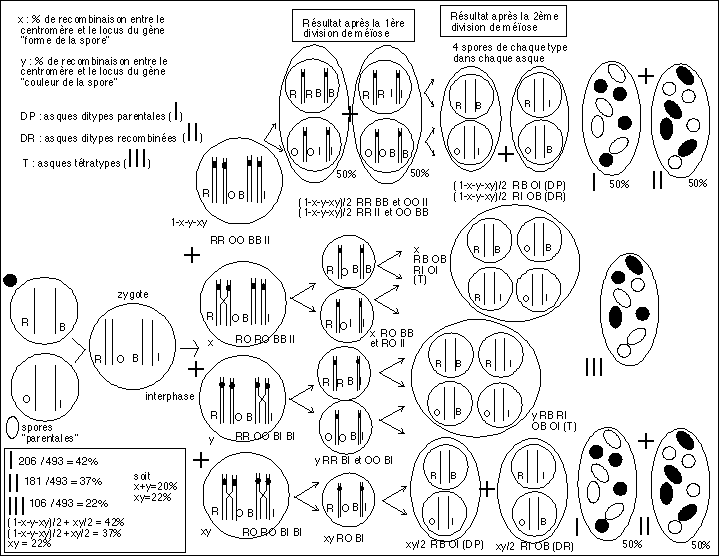
On peut essayer de suivre le même raisonnement avec deux caractères : je prend l'exemple d'un exercice classique de TC (manuel scolaire Hachette, édition de 1984, p80) avec un champignon "haploïde" : Ascobolus immersus dont les octades au sein des asques ne sont pas ordonnées. Je vous propose ici une interprétation SANS MEIOSE NI CROSSING-OVER... on a deux couples de caractères "couleur de la spore" : brune (B) ou incolore (I) et "forme de la spore" : ronde (R) ou ovale (O). On sélectionne des spores rondes et brunes et des spores ovales et incolores que l'on fait pousser. On obtient une fécondation (zygospore) puis la formation d'asques contenant des spores (ascospores) qui se répartissent de la façon suivante : 206 asques présentent 4 spores rondes et brunes et 4 spores ovales et incolores (asques que l'on appelle classiquement ditypes parentales), 181 asques possèdent 4 spores ovales et brunes et 4 spores rondes et incolores (asques nommées ditypes recombinées), enfin 106 asques ont 2 spores rondes et brunes, 2 spores rondes et incolores, 2 spores ovales et incolores et 2 spores ovales et brunes (tétratypes).
Interprétation classique avec 2 gènes indépendants, 4 allèles, une méiose et des crossing-over..
Voici une interprétation non conventionnelle SANS MEIOSE donc avec une phase de maturation sexuelle qui aboutit à un accolement des homologues qu'ils soient bi ou monochromatidiens, et qui ne se séparent qu'à l'anaphase de la mitose qui suit. Il n'y a PAS NON PLUS DE CROSSING-OVER mais simplement une mitose qui précède ou suit la phase de maturation sexuelle.
Interprétation NON CONFORMISTE sans méiose ni crossing-over....