|
Cette petite page n'a
pas la prétention de "traiter" ces délicates
questions dans leur ensemble. Elles font l'objet
d'innombrables traités érudits. J'y ai juste
précisé quelques points qui me paraissent
nécessaires pour aborder ces questions avec ouverture
d'esprit.
|
||||||
|
Plan |
|
|||||
|
|
* en biologie on nomme les espèces animales (voir partie 2) alors que les hommes se nomment comme individus (voir le chapeau d'anthropologie sur la page de 1èreES ou 1èreL sur la lignée humaine). |
|||||
|
Une nomenclature
|
La nomenclature des espèces vivantes désigne donc plus spécifiquement les noms des espèces vivantes. |
|||||
|
Le premier point, qui peut surprendre,
est que le nom d'espèce ne se donne pas seul : il est
toujours précédé par un nom de genre.
Toute individu à donc deux noms (c'est donc une
dénomination que l'on qualifie en latin de
bi-nominale = deux-noms) inséparables
pour désigner son espèce. |
Pour la zoologie: Pour la botanique: * taxon: élément d'une classification (la taxinomie est la science des classifications). |
|||||
|
Nom
d'espèce complet de l'homme
: Animal,
Vertébré, Mammifère, Primate,
Hominidé, Homo sapiens? |
|
|||||
|
|
Certaines personnes tentent de promouvoir une nomenclature non-linnéenne, sans hiérarchisation des regroupements de taxons, pour assurer une ressemblance superficielle avec la méthode cladiste : le phylocode. |
|||||
|
|
Il est courant que l'on identifie trop
rapidement les termes genre ou espèce
des philosophies, comme celle d'Aristote, avec les
catégories classificatoires linnéennes. Si les
liens sont manifestes (du fait historique du choix des
termes par Linné), il faut se méfier d'une
lecture trop rapide: l'espèce de Thom
ou d'Aristote (eidos) est plutôt, pour un
esprit moderne, la forme. René Thom
emprunte à Aristote le Genre, (que l'on peut
se représenter comme un arbre infini qui à
partir d'un individu donne une infinité de
descendants... et qui ressemble à un continu : selon
les mots d'Aristote : «Genos hos
hylè» : le genre est «comme une
matière»). Il l'oppose à l'espèce
(eidos) , sous-division du genre (Thom la compare
à une sous-variété -
mathématique: si le genre est considéré
comme un espace euclidien; «en codimension 1 elle sera
définie comme le lieu d'une fonction locale
régulière appelée par Aristote la
différence spécifique»).
|
Références Œuvres
complètes de René Thom (CD-Rom IEHS) |
||||
|
|
Une classification est une répartition en classes, groupes ou catégories d'éléments partageant un ou plusieurs critères. |
|||||
|
Classification naturelle objective ou classification artificielle subjective |
Avec Linné et Buffon deux visions
s'affrontent: Colloque : Les mots et les
choses au XVIIIe siècle : la science, "langue bien
faite" ? |
|
||||
|
La distinction faite ici entre
continuum et discrétisation des
espèces, me semble différente de ce que
certains épistémologues et historiens des
sciences (voir par exemple l'article "Classification du
vivant" dans l'EU2009 par Pascal Duris et Pascal Tassy)
croient pouvoir présenter comme une opposition entre
une "nature continue" (qu'ils rapportent paradoxalement
à l'image d'une échelle) et une "nature
discontinue" (qu'ils rapportent douteusement à une
conception plus ou moins créationniste). Pour moi il
est clair que c'est la continuité
de l'espèce qui est en jeu
et non le concept de nature (voir ci-dessous).
La nature n'est pas un continu, que ce soit au niveau
moléculaire, individuel ou encore populationnel....
il y a plein de niveaux de discontinuité en elle. |
||||||
|
Toute classification phylogénétique est naturelle |
Depuis l'avènement de l'idée d'évolution on peut penser que le problème est résolu. Que se soit avec la vision vitaliste de Lamarck ou celle de l'évolutionnisme socio-politico-biologique de Darwin, les classifications des espèces vivantes sont désormais phylogénétiques, c'est-à-dire qu'elles s'efforcent de rendre compte de l'évolution des espèces (et donc des liens de parenté entre elles). La phylogénie (de phylum = la lignée), de science du développement et de l'évolution des êtres vivants, est devenu pour beaucoup simplement l'histoire généalogique de la vie. Dans ce cas la phylogénie est une et c'est un travail d'historien que doit faire le paléontologue (même si ce travail se fait avec un tout autre type d'archives que celles de l'histoire). L'évolution a donc ainsi résolu le problème de la pertinence d'un ordre classificatoire puisqu'il suffit de représenter un ordre historique. Tout le monde peut donc s'accorder et dire qu'il existe une classification naturelle qui est phylogénétique. Mais c'était sans compter sur les inattendus de l'histoire. Par un tour de passe passe comme l'histoire en recèle tant d'autres, certains en sont revenus à nier la présence d'un ordre naturel en réintroduisant le hasard comme mécanisme de l'histoire du vivant principalement dans la théorie synthétique de l'évolution.
|
Pour une approche érudite de l'histoire des classifications jusqu'au début du XXème,, je conseille un texte d'André Pichot: Définir, décrire et classer en biologie, Delagrave 2005 |
||||
|
Le sens de l'histoire |
Sans prétendre trancher cette
question, je crois que l'on peut affirmer que les luttes ont
continué à cause d'un profond désaccord
sur le sens de l'histoire.
|
|||||
|
|
Depuis l'avènement de
l'idée
d'évolution
la classification des êtres vivants ne classe
plus des INDIVIDUS mais des ESPÈCES, car ce
n'est pas l'individu qui évolue mais les
espèces qui se transforment. Il est donc
préférable de parler de la
classification des espèces.
|
 Le titre de l'ouvrage de Darwin annonce une théorie de l'origine des espèces alors qu'il traite principalement de leur disparition au moyen de la sélection naturelle. |
||||
|
|
||||||
|
La taxinomie |
La science de la classification des êtres vivants est la systématique ou la taxinomie. |
On parle aussi de biodiversité pour signifier la diversité des êtres vivants mais on a tendance à élargir cette notion à l'ethnodiversité (diversité des hommes) et plus récemment à la géodiversité (diversité des roches, minéraux, paysages et autres formes physiques terrestres...). |
||||
|
|
||||||
|
3.1 L'étape cladiste
|
||||||
|
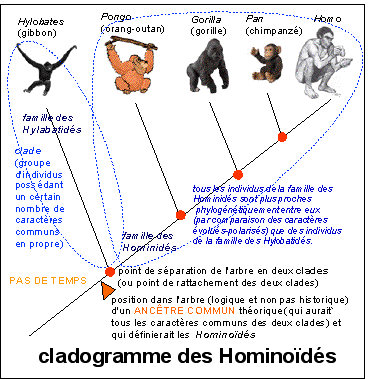
La méthode cladiste ou cladistique
construit, selon une méthode élaborée
par Willi
Hennig (Grundzüge einer
Theorie der phylogenetischen Systematik ,
W. Hennig, 1950), des
cladogrammes
qui traduisent graphiquement les
clades
ou lignées évolutives
construites à partir de caractères
polarisés (dont on décide s'ils sont
primitifs ou évolués à l'aide
d'hypothèses évolutives
paléontologiques et embryologiques).
|
Voir quelques éléments pour comprendre les classifications cladistes ci-dessous. |
|||||
|
Un cladisme ouvert à plusieurs théories évolutives |
Le cladisme part des caractères
(des organismes actuels et des fossiles) et s'efforce de les
relier phylogénétiquement avec le plus de
rigueur logique possible. Il suppose l'évolution mais
ne favorise aucune théorie
particulière. Toute théorie
évolutive peut servir à polariser les
caractères (voir
page sur le
cladisme).
|
|||||
|
Il n'y a pas de temps pour le cladiste
Voir cours de seconde
: |
Si les phylogénies construites à partir d'analyses cladistiques sont maintenant la norme, il ne faut pas cependant pas croire que c'est la méthode qui fournit l'histoire. Il n'y a pas de temps pour le cladiste, mais seulement des relations logiques de parenté. |
Mais bien évidemment, le paléontologue qui cherche à reconstituer l'histoire du vivant utilise les datations réalisées à partir des fossiles (par d'autres méthodes que le cladisme : datation logique ou datation expérimentale). L'histoire vient des fossiles que l'on plaque sur le cladogramme et qui devient ainsi un arbre phylogénétique. En ajoutant ainsi le temps on sort du cladisme pour rentrer dans l'histoire du vivant. Mais cette construction pose bien des problèmes car comment concilier une histoire, forcément unique, avec une construction logique parcimonieuse, forcement plurielle. |
Ainsi, avec la cladistique on a progressé en scientificité car la méthode est unifiée et les critères sont clairs, mais on a aussi régressé en terme de compréhension car ce n'est pas le cladisme qui peut permettre de reconstituer l'histoire (mais ce sont bien les fossiles et leur interprétation).
|
|||
|
le cladisme peut facilement être dévié vers un relativisme* philosophique |
Sous des apparences consensuelles le relativisme (en morale un relativiste pourrait s'exprimer ainsi "tout n'est qu'opinion et toutes les opinions se valent") est extrêmement violent vis-à-vis de ceux qui le rejettent. Associé au scientisme c'est une idéologie tyrannique (voir Qu'est-ce que la science ?). La ressemblance entre deux
individus A et B plus proches entre eux que d'un autre C
(similitude relative de A et B par rapport à C)
s'exprime alors par l'idée que leur ancêtre
commun le plus ancien logiquement est plus récent
logiquement que l'ancêtre commun théorique
qu'ils ont chacun ou à deux avec cet individu C.
C'est cette notion de ressemblance relative qui me
paraît fondamentalement différente d'une
classification qui cherche à établir des
parentés absolues (historiques), entre deux
groupes. |
Remarque: |
||||
|
Pour un enseignant le foisonnement des classifications cladistes plus ou moins signifiantes est une difficulté supplémentaire, sans parler des nouvelles dénominations des groupes qui, par définition, ne seront jamais unifiées mais resteront plurielles. Il est patent que la méthode qui devait permettre de connaître enfin L'histoire de la vie, n'a pu donner naissance qu'à des historiettes : c'est le tribut à payer au relativisme. Mais on peut réagir. |
||||||
|
Les données moléculaires, et leur traitement statistique, ont d'abord permis un développement des méthodes phénétiques (basées sur les ressemblances sans s'occuper des relations de parenté exprimées au niveau des caractères). Ces données moléculaires se sont ensuite avérées être facilement utilisables en cladistique si l'on faisait des hypothèses simples de relations évolutives entre molécules, ce que fournissait la théorie de l'information génétique (des distances exprimées en nombre minimal de substitutions et autres "mutations"). |
Mais, désormais, les outils mathématiques utilisés par la cladistique -ou la phénétique- lorsqu'elle compare des caractères se sont grandement sophistiqués. En gagnant toutes les écoles de classification les méthodes statistiques ont véhiculé avec elles, de façon parfois cachée, le réductionnisme génétique (théorie de l'information génétique), qui renaît ainsi de ses cendres. L'utilisation des statistiques pour l'analyse de caractères morphologiques pourrait évacuer ce problème (voir l'exemple de la lignée humaine), mais en pose d'autres à son tour. |
Il y a un vrai paradoxe: d'un côté on a une biologie moléculaire de plus en plus probabiliste (de moins en moins déterministe) et de l'autre des mécanismes moléculaires évolutifs dépassés qui servent à évaluer statistiquement des lignées évolutives. Et si l'on veut essayer de comprendre la valeur historique de ces nouveaux arbres phylétiques on est forcé de constater qu'elle repose sur des éléments disparates qui ne possèdent pas le même degré de confiance (il s'agit ici de la confiance que l'on fait à toute preuve scientifique) : horloge moléculaire (encore fort utilisée, notamment pour les branchements), datation des fossiles, hypothèses anatomiques ou embryologiques (la formation de telle structure ne pouvant intervenir que postérieurement à telle autre)... |
||||
|
Conclusion de l'article "Classification
du vivant" de Duris et Tassy in EU2009 « La conclusion de cette brève histoire des classifications nous invite à une humilité qui est désormais aux antipodes des standards des publications scientifiques. Si un siècle après L'Origine des espèces, on a admis que les classifications devaient être phylogénétiques, on s'est vite désaccordé sur les méthodes permettant de reconstruire la phylogénie. Cet aspect n'apparaît pas aux yeux du grand public : un résultat phylogénétique est le plus souvent présenté comme un résultat biologique qui va de soi, s'imposant de lui même, où la méthode de travail n'est pas censée influencer le résultat. Ce n'est pas tout à fait vrai. Alors que la classification a d'abord été vue comme un système de référence invitant à la stabilité du savoir, la phylogénétique est un domaine en recherche » |
||||||
|
|
||||||
|
Enseigner un cladisme sans relativisme et une histoire du vivant |
|
|||||
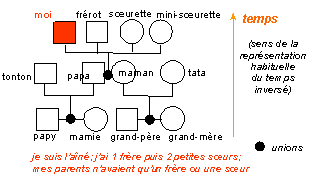
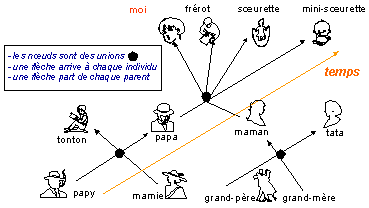
|
Une
généalogie... |
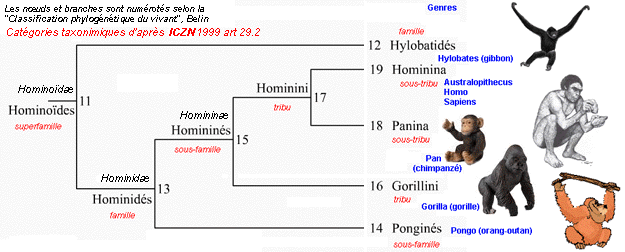
un cladogramme où les nœuds représentent la position logique des "ancêtres communs" qui sont des entités théoriques regroupant les caractères de plusieurs clades
voir cladogramme
complet du
Cours
1ES-1L (partie 2.1) |
Les enseignants,
et majoritairement les enseignants du
secondaire, utilisent la plupart du
temps des
arbres
phylogénétiques
(qui ne sont donc plus des
cladogrammes)
à des fins pédagogiques pour
représenter l'évolution. Ce faisant l'enseignant se heurte alors
au réalisme "naturel" des élèves qui
tentent par tous les moyens de plaquer l'histoire
(qu'ils savent être unique puisque le temps est une
caractéristique de la vie) sur ces cladogrammes. Ce sont les élèves qui ont raison. En effet, pour être juste, il ne
faut pas mélanger histoire et cladogramme. |
||||
|
|
||||||
|
La vie comme continu: |
Il existe d'autres pistes pour
éviter les confusions histoire-cladisme (ou
plutôt généalogies-cladogrammes) dans
l'esprit des élèves. |
|||||
|
Addenda à l'attention des collègues enseignants: |
||||||
|
hasard, contingence, darwinisme, scientisme, déterminisme, créationnisme «...la science
enfle...» |
Les théories de l'évolution ont une dimension philosophique et politique qu'il est vain d'essayer d'occulter. La dimension religieuse n'est pas plus marquée que pour toutes les autres questions de la biologie qui touchent à l'homme (la question de l'anthropologie est abordée dans la page sur l'évolution de l'homme). |
On rapporte (voir ci-contre
"Un autre Darwin" <---) des prises de positions
anti-évolutionnistes inacceptables de certains
élèves qui, très secondairement, se
prétendent créationnistes, mais qui sont
plutôt dans une démarche intégriste
voire sectaire... |
||||
|
|
||||||
|
«Les principes éthiques forment le socle de la science [... ] . La recherche de la vérité et l'idée d'une approximation de la vérité sont aussi des principes éthiques; tout comme le sont les notions d'intégrité intellectuelle et de faillibilité, qui conduisent à une attitude d'auto-critique et à la tolérance.» Karl Popper (citation en haut de la page d'accueil de ce site) |
Page sur le darwinisme moderne extrait de l'Histoire de la notion de vie, André Pichot, Gallimard, 1993 |
|||||
|
Je crois
qu'il est du ressort de l'Éducation Nationale de
replacer correctement le débat en montrant les
positions philosophiques de chacun. Un enseignant doit
pouvoir enseigner qu'il existe des courants non darwiniens
en biologie et en paléontologie. Un enseignant
croyant doit pouvoir affirmer, contre l'avis d'athées
militants comme G. Lecointre, qu'il n'y a pas
d'incompatibilité entre science et foi et qu'au
contraire l'une et l'autre grandissent l'homme et que l'on
peut tous travailler à la
vérité.
|
||||||
|
Enseigner l'évolution avec ouverture d'esprit |
On comprend le souhait d'harmonisation des instances pédagogiques de l'Éducation Nationale mais harmonisation ne veut pas dire homogénéisation. C'est ainsi que l'on a favorisé des positions sectaires. |
J'encourage donc les collègues
à enseigner plusieurs théories de
l'évolution, à ne pas cacher leur foi ou leur
athéisme sans en faire l'ostentation et à
toujours accepter le débat. |
||||
|
|
||||||
|
Pour une approche érudite de l'histoire des classifications jusqu'au début du XXème,, je conseille un texte d'André Pichot: Définir, décrire et classer en biologie, Delagrave 2005 |
Je veux aussi soulever ici le problème de l'usage immodéré de l'ouvrage de Le Guyader-Lecointre: Classification phylogénétique du vivant (Belin). Dans cet ouvrage la présentation
de ce qu'est une classification est extrêmement
faussée (p11-18). |
*Il n'est pas
acceptable que l'Éducation Nationale se fasse le
relais d'une philosophie relativiste sans présenter
d'autres approches: |
||||



|
page plus détaillée sur le cladisme |
||||||
|
en
cladistique, il n'y a pas de temps, il n'y a que des
relations logiques
|
||||||
|
comparer des similitudes = deux caractères qui se ressemblent fortement |
soit homologie* (véritable caractère hérité) |
présence de vertèbres (même si toutes les vertèbres ne sont pas identiques on peut considérer que le fait de posséder des vertèbres est un caractère homologue chez deux individus, même éloignés). Tous les animaux à vertèbres** forment donc un clade (groupe monophylétique: ayant une origine commune; le point de rencontre de tous les caractères communs aux membres du clade est appelé "ancêtre" commun). |
* l'homologie et l'analogie cladistes n'ont pas le sens classique, voir page sur le cladisme ** la grande importance de ce caractère pour classer les animaux a été reconnue dès la fin du 18ème siècle; on attribua par exemple à Geoffroy Saint-Hilaire la chaire des Animaux vertébrés lors de la création du Muséum (1793); Lamarck du se contenter de la chaire des insectes et vers (voir discours de 1801 (p7); dans une toute autre démarche classificatoire, avec son Systema Naturæ, Linné utilisait des caractères moins anatomiques et plus physiologiques...(REGNUM ANIMALE extrait de la 12ème édition de Systema naturae, 1766, Gallica-BNF, tome 1 (partie 1) p 19) |
|||
|
|
||||||
|
soit analogie* (caractère semblable par convergence ou hasard mais sans lien de parenté direct évident) |
présence
d'écailles : toutes les
écailles n'ont pas forcément la même
structure ni la même origine (les écailles des
poissons ne sont pas homologues des écailles des
reptiles ou des oiseaux)
|
ancienne page de terminale sur les plumes |
||||
|
|
||||||
|
comparer des homologies = caractères hérités |
soit caractère à l'état dérivé (est l'expression d'une parenté = résulte de l'évolution); noté 1 |
synapomorphie (partage de l'état apomorphe (du grec apo = loin de et de morphos = la forme) = caractères éloignés |
la présence
d'une phanère de type "
plume" de l'Archéoptéryx est un
caractère dérivé par rapport à
l'écaille d'un reptile ancestral qui serait d'une
lignée n'ayant jamais eu de représentants
à plumes (par
exemple
Sinosauropteryx);
le groupe des
reptiles comportant des plumes (Archæopteryx,
Confuciusornis, Enantiornithines,
Euornithes)
et qui aurait donné naissance aux oiseaux serait ce
que l'on appelle un groupe monophylétique (qui
contient un ancêtre commun et tous ses "descendants").
|
|||
|
|
||||||
|
soit caractère à l'état primitif ou à l'état ancestral (est le résultat d'une stabilité); noté 0 |
plésiomorphie (du grec plésios = voisin et morphos = la forme) = caractères voisins |
la présence de
plumes chez deux oiseaux est un
caractère primitif (voisin)
|
||||
|
|
||||||
|
"l'ancêtre commun" et les "descendants" de l'ancêtre commun (formant un groupe monophylétique = clade) |
le terme d'ancêtre ne désigne pas un fossile mais un concept (ce n'est pas un individu) qui regroupe un certain nombre de caractères. Quand on parle d'ancêtre "antérieur", il faut comprendre "plus éloigné en terme de similitude". Un ancêtre commun à deux groupes-frères est le point de rencontre logique le plus proche de tous les caractères propres à chaque groupe. Ce que le paléontologue interprète comme un indication bien sûr de l'emplacement LOGIQUE du véritable ancêtre (mais hypothétique), historique, d'une lignée qui posséderaient les caractères portés par le clade. |
|||||
|
|
||||||
|
Extraits de et commentaires sur l'article "Classification du vivant" dans l'Encyclopedia Universalis de 2009, signé par Pascal Duris (Professeur d'épistémologie et histoire des sciences à l'Université Bordeaux 1) et Pascal Tassy (Professeur au Muséum d'Histoire Naturelle) |
||||||
|
Sans constituer encore une rupture, cet article de vulgarisation marque une nouvelle étape après la conquête de l'enseignement par un certain cladisme: la reconnaissance de l'existence de différents points de vue. |
Je suis particulièrement agacé par l'utilisation dans un sens volontairement(?) falsifié (scientiste si je comprends bien) de certains mots comme " créationniste" ou "chronologie biblique". Je suis las de cette "mauvaise foi", mais c'est sans doute un problème récurrent en histoire. |
Une fois encore ce n'est pas dans leur présentation de l'histoire ancienne des classifications que ces auteurs sont crédibles mais dans les quelques éléments de l'histoire récente qu'ils signalent. |
||||
|
* somme des
modifications |
Une clé de l'argumentaire des
auteurs repose sur l'idée que Darwin aurait
considéré, dans son effort de reconstitution
de la généalogie des êtres vivants, une
sorte de
«dualisme»
pour les caractères qui seraient soit une
«somme des modifications» soit des
«indications de descendance». Duris et
Tassy soulignent ainsi, dans l'histoire récente, une
préoccupation excessive pour les caractères
vus comme la "somme des modifications" (dans le cladisme),
alors que persiste la difficulté de retrouver des
"indications de descendance". |
|||||
|
Charles Darwin, L'origine des espèces au moyen de la sélection naturelle ou La lutte pour l'existence dans la nature (1859), d'après l'édition de 1896 (Ed. Schleicher frère), traduit de l'édition anglaise définitive par Ed. Barbier (accessible sur le site de l'ABU ou mieux, celui de l'Université du Québec à Chicoutimi: http://classiques.uqac.ca /classiques /darwin_charles_robert /origine_especes /origine_especes.html )
|
|
|||||
|
La somme des modifications c'est
l'étendue des différences. |
||||||
|
Charles Darwin, The descent of man, and selection in relation to sex, London, John Murray, 1st edition, 1871 sur le site Darwin_online: http://darwin-online.org.uk /EditorialIntroductions /Freeman_TheDescentofMan.html et traduction d'Edmond Barbier, 1891. La descendance de l'homme et la sélection sexuelle. Preface by Carl Vogt. Paris: C. Reinwald à droite, sur le même site |
|
|
||||
|
|
||||||
|
Les auteurs soulignent aussi le délicat usage des grades (niveaux évolutifs). Mais il me semble que si l'on affirme que des organismes, même s'ils n'appartiennent pas à la même lignée, appartiennent à un même grade s'ils sont au même stade évolutif, cela sous-entend l'existence d'un mécanisme évolutif identique. En effet, il ne s'agit pas ici de datation, mais bien de complexité, ou, au moins, de niveau dans un processus évolutif supposé unique.. |
De plus, ces auteurs rappellent (mais est-ce vraiment un "rappel" ?) «les concepts formalisés en 1870 par le zoologiste britannique Edwin Ray Lankester qui décomposa la notion d'homologie. Les homologies dues à la parenté sont appelées « homogénies » ; les autres, présentes chez diverses espèces sans pour autant avoir été héritées d'un ancêtre commun, sont appelées « homoplasies ». Depuis, on a restreint le sens de l'homologie à celui de l'homogénie, ce dernier terme tombant dans l'oubli.» |
|||||
|
|
||||||
|
irruption des statistiques |
Enfin, le point majeur, à mon
sens, est la présentation qu'ils font de l'irruption
des statistiques dans la classification. |
|||||
|
«L'ordinateur, la molécule et
la ressemblance
La systématique cladistique, en se fondant sur la notion d'homologie, se trouve alors à la fois être dans la droite ligne des idées darwiniennes et offrir une mathématisation nouvelle parfaitement adaptée au traitement informatique, alors en pleine émergence. À cette époque, les systématiciens ont les moyens techniques d'analyser un nouveau type de caractères : les caractères génétiques. Ils prennent en compte la structure des protéines, puis celles de l'ADN et de l'ARN. Ces caractères, dits discrets (discontinus), se conforment apparemment bien à la notion d'homologie. La comparaison des séquences permet d'émettre des hypothèses de transformations d'états : tel acide aminé se substituant à tel autre, tel nucléotide à tel autre. Dans les années 1980, avec l'essor des ordinateurs personnels, la construction des arbres phylogénétiques utilisant des caractères morphologiques et moléculaires, ou bien séparément ou bien simultanément, se multiplie au point de devenir routinière. Très vite, pourtant, de nouveaux algorithmes de regroupement, non parcimonieux, sont mis au point et consacrent la fin de l'alliance méthodologique un temps entrevue par la cladistique. Cependant, avant d'aborder cette dernière étape de l'histoire des classifications, il convient d'évoquer l'autre révolution en matière de systématique : la « révolution phénétique » ou l'école de la taxinomie numérique. En effet, dans les années 1960, des mathématiciens intéressés par les classifications biologiques et des biologistes fortement nourris de statistiques proposèrent d'abandonner la recherche des homologies pour se consacrer au calcul du degré de ressemblance afin de regrouper les espèces. Inventant un nouveau langage et de nouvelles méthodes, proposant des algorithmes de calculs des ressemblances, ils firent référence, à l'instar de l'Américain Robert Sokal, à l'objectivité des mathématiques. L'indice de ressemblance calculé entre deux espèces (unités taxinomiques opérationnelles ou U.T.O.) ne devrait pas souffrir d'ambiguïté. De la sorte, les arbres de parenté construits par les phénéticiens, appelés phénogrammes, n'ont pas vocation à raconter l'histoire de l'évolution et n'expriment que la ressemblance toutes catégories confondues (qu'il s'agisse d'états primitifs, d'états dérivés ou encore d'homoplasies). Cependant, très vite, ce qui était au départ une nouvelle école de classification devint surtout une école de constructions d'arbres. Et l'on s'aperçut qu'avec les mêmes données, différents calculs d'indices de ressemblance pouvaient donner des groupements différents, ce qui marqua la fin de l'objectivité mathématique. Il n'en reste pas moins vrai que les statisticiens n'arrêtèrent pas de proposer de nouveaux calculs, fournissant ainsi aux systématiciens des logiciels efficaces de constructions d'arbres de similitude, tous baptisés arbres phylogénétiques. |
(suite)
On imagine la fureur des
systématiciens évolutionnistes devant ce
qu'ils considéraient comme une récession.
Pourtant, la phénétique entraîna de
nombreux phylogénéticiens : il suffit
d'admettre a priori que le mélange des trois
catégories de ressemblance et la fusion de
l'homologie et de l'homoplasie ne présentent pas trop
d'inconvénients &endash; s'annulent en quelque sorte
&endash; pour considérer qu'un schéma de
ressemblance globale peut être un schéma
phylogénétique. Comme en cladistique, les
données moléculaires furent donc
intégrées dans les approches
phénétiques dès la fin des
années 1960.
Il n'est pas besoin de préciser que la critique mutuelle que se font parfois les partisans des méthodes probabilistes et les partisans des méthodes cladistiques peut se révéler aussi féroce que lors des débats des années 1970 entre les évolutionnistes, les cladistes et les phénéticiens. Certains considèrent que les résultats tirés des approches probabilistes sont un nuage de fumée, d'autres que la morphologie n'est pas fiable et a fait son temps. L'enjeu est toujours le même : imposer un système de référence général, ou tout au moins installer au premier plan sa propre école méthodologique. En outre, les groupements d'espèces obtenus par ces méthodes sont formalisés comme l'est tout groupement d'espèces obtenu par traitement de l'homologie et de l'homoplasie. Des groupes impossibles à distinguer à l'aide de caractères morphologiques sont obtenus au moyen de caractères moléculaires traités sur le mode probabiliste. Aussi, pour terminer par des exemples empruntés aux mammifères, citera t-on un succès de la biologie moléculaire avec l'identification du groupe Cetartiodactyla, incluant dans la nature actuelle les Cetacea (baleines et cachalots) et les Artiodactyla (cochons, hippopotames, chameaux et ruminants), voire la proche parenté entre les baleines et les hippopotames. À l'inverse, un groupe comme les Afrotheria (qui inclut notamment des formes aussi distinctes que les éléphants, les macroscélides et les taupes dorées), obtenu par l'analyse probabiliste de différents gènes, est actuellement une sévère pomme de discorde entre les systématiciens morphologistes et molécularistes. Aucun état dérivé de traits anatomiques ne permet pour l'instant d'associer ces différents mammifères. En outre, les horloges moléculaires estiment que la différenciation non seulement des Afrotheria mais aussi des grands groupes de mammifères placentaires aurait été pleinement réalisée dès le Crétacé, bien avant que les premiers représentants desdits groupes soient connus.» |
|||||

{kind=link}
{kind=link}