|
retour plan du cours, accueil, panorama |
|
Quelques
références: |
||||||
|
Nouvelle version en travaux juillet 2009 les ouvrages de vulgarisation sur le cladisme n'ont pas suivi les développements rapides dans les laboratoires. Je livre ici ce qu'en tant qu'enseignant je m'efforce de comprendre. je serai honoré d'avoir d'autres ouvertures. |
|
|||||||
|
Plan
|
|
|
||||||
|
|
|
|||||||
|
1. Le petit cladiste illustré: comprendre comment passer d'une classification d'objets à une classification évolutive Au début, c'est très simple, on voit bien qu'il ne s'agit que de comparer deux objets... on voit clairement que l'on est dans un discours logique et non pas naturaliste (biologique). |
|
Avertissement: |
||||||
|
On classe des objets logiquement sans se préoccuper de l'histoire |
|
|
||||||
|
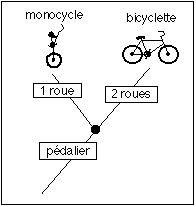
Un exemple de cladogramme simple avec deux objets à roues permettant de s'asseoir...
Ce qui compte c'est la distribution des caractères pour chaque branche ou groupe de branches (réunies au niveau d'un nœud, représenté par un point noir)
|
|
Les
éléments
de comparaison
qui sont
choisis comme
déterminants
sont
encadrés.
On les nomme caractères.
On voit bien
dans le cas
présent
que ce
cladogramme ne
représente
pas une
histoire mais
juste une comparaison;
un
cladogramme est
un «graphe
connexe non
cyclique»
disent des
cladistes (Classification
phylogénétique
du vivant,
G. Lecointre
et H. Le
Guyader, 2001,
Belin, p 21).
le
nombre de
roues (et
donc le point
par où
passe le
vecteur poids
qui part du
centre de
gravité
et est
dirigé
vers le centre
de la terre) est
dérivé
(logiquement
et non
historiquement;
voir par
exemple http://pages.
globetrotter.net/
groleau/ amq/
francais/
history.htm
) par rapport
à
l'invention de
la roue (?) et
la la position
du
pédalier
horizontale ou
verticale est
aussi dérivée
de la
présence
d'un
pédalier
(transmission
d'une force
verticale en
rotation) ,
que celui-ci
soit
centré
sur la roue ou
décalé
grâce
à
l'intervention
d'un axe
supplémentaire
et d'un moyen
de
transmission.
On a donc
choisi de
faire un clade
avec les
véhicules
à
pédalier
et roue(s) et
selle (par
exemple). Tous
ceux qui n'ont
pas ces 3
éléments
ne sont pas
dans le clade
(outgroup). Le
nombre de
roues est ici
considéré
comme une
caractère
innovant, une
synapomorphie
(voir-ci-dessous)
avec 2
valeurs: 1 ou
2 roues. On
peut aussi
choisir comme
caractère
dérivé
le
décentrage
horizontal ou
vertical du
pédalier
par rapport
à l'axe
de la roue et
au sol. |
||||||
|
Commentaire L'évolution
comme
bricolage |
|
|||||||
|
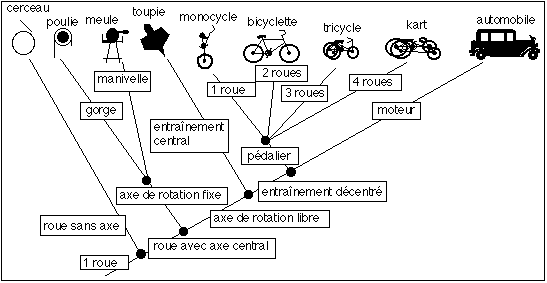
Mais en ajoutant des objets et des caractères, tout se complique et le temps réapparaît comme par magie... toujours de façon erronée, mais le dépistage de l'erreur devient nettement plus difficile: |
|
|
||||||
|
|
|
l'évolution
se fait dans
l'ordre
suivant (il
s'agit d'un
ordre logique
et non
historique);
ces inventions
dérivant
logiquement
les unes des
autres (mais
pouvant
très
bien
apparaître
dans un ordre
temporel
différent
(ceci
est par
exemple
évident
pour le nombre
de roues ou
l'invention
d'une toupie
par rapport
à une
meule ou un
cerceau....)):
roue sans axe
et avec
déplacement,
roue avec axe
fixe, axe avec
mouvement
d'entraînement
décalé
/ avec axe
centré,
axe et
déplacement,
point
d'entraînement
très
proche du
centre ou
décentré,
force
mécanique
musculaire /
thermique-électrique,
1-2-3-4 roues groupe monophylétique = clade = branche (clade) ou ensemble de branches (groupe) de l'arbre qui contient un ancêtre et tous ses descendants (la branche "automobile" forme un clade à elle toute seule, comme l'ensemble monocycle, bicyclette, tricycle et kart) |
||||||
|
|
groupe polyphylétique = groupe qui n'a pas d'ancêtre commun (qui contient au moins une homoplasie) par exemple, dans notre représentation, la toupie forme un groupe polyphylétique avec la poulie et la meule innovation 1 = synapomorphie = caractère dérivé = caractère (à l'état) apomorphe; caractère qui provient d'un autre avec modification: deux synapomorphies sont séparées par un nœud; la manivelle est un caractère dérivé de l'axe de rotation fixe ; en fait un caractère dérivé est une relation logique de similitude, il n'entre dans cette relation aucune idée de temps. innovation 2 = plésiomorphie = caractère primitif = caractère ancestral = caractère (à l'état) primitif ; caractère "non évolué" ou original que l'on ne trouve pas dans un autre groupe apparenté (une plésiomorphie peut être partagée par plusieurs taxons: c'est une symplésiomorphie); ici c'est la roue qui était le caractère primitif ; en fait un caractère primitif est une relation logique de similitude (il n'y a aucun élément voisin semblable qui possède le même caractère de façon aussi originale - c'est-à-dire sans lien avec d'autres caractères, logiquement associés et qualifiés de dérivés - c'est une relation de dissimilitude et non d'antériorité), il n'y a encore ici aucune idée de temps. |
|||||||
|
|
|
homoplasie = similitude entre deux taxons (éléments de la classification) qui ne dérive pas d'une relation de parenté et donc d'un ancêtre commun - par exemple le cerceau et une poulie; mais aussi , dans notre choix de représentation, la présence de plus d'une roue entre la bicyclette et l'automobile (alors quec'est une homologie de filiation entre la bicyclette et le tricycle ou le kart.... toujours selon nos choix de représentation : on voit facilement que l'on pourrait faire un choix tout à fait différent en considérant le nombre de roues comme une homologie pour tous les taxons...). convergence = cas particulier d'homoplasie mais répétée chez plusieurs taxons (le présence de 4 ou 3 roues pour tous les véhicules à moteur pourrait être considéré comme une convergence) - pour les non-cladistes cette similitude était considérée comme résultant d'un même facteur, le plus souvent l'action du milieu... analogie cladiste = cas particulier d'homoplasie où les deux caractères ont la même fonction: le cerceau et la roue de vélo par exemple |
|
|||||
|
|
|
phylogénie cladiste = classification évolutive une classification cladiste ne reflète pas l'histoire: un arbre cladistique n'est pas une généalogie; l'évolution, pour le cladiste, n'est pas dans l'arbre mais dans la méthode (polarisation des caractères principalement); dans notre exemple, une classification historique se préoccuperait de savoir quels ont été les brevets déposés, quelles ont été les conditions sociales, culturelles, politiques de l'avancée des inventions... alors que l'arbre cladiste ne reflète que la considération actuelle des inventions, qu'il projette en arrière dans le temps. |
|
c'est pour cela que les classifications cladistes ne sont pas vraiment évolutives dans le sens où elles n'en apportent pas la preuve (au sens d'argument) mais elles illustrent une évolution qui est ailleurs. Les classifications vraiment évolutives sont des classifications qui prennent l'évolution comme causalité et cherchent à en préciser les modalités. Elles donnent un sens biologique à l'évolution: perfectionnement, diversification, complexification, sommation et mise en mémoire... que l'on y mette un vitalisme ou un matérialisme ou toute autre philosophie. Il y a une séduction trompeuse dans le travail de classification cladistique : au fur et à mesure que l'on accumule les caractères, en décidant progressivement de leur caractère primitif ou dérivé, on se forge une histoire, que l'on en vient progressivement à considérer comme l'histoire. On ne peut s'empêcher de placer le temps verticalement alors que dans la méthode il n'y a PAS de TEMPS. Il n'y a que des comparaisons. |
||||
|
|
|
ancêtre commun l'ancêtre commun n'existe pas réellement, il n'a pas existé; le premier objet à roues n'était ni un monocycle ni une bicyclette, il possédait seulement une roue; le rechercher relève de l'impossibilité historique. Les ancêtres communs NE sont PAS aux NŒUDS de l'arbre (qui séparent simplement les synapomorphies), mais à leur voisinage. |
|
Cette notion cladiste de parenté réduite à l'ancêtre commun - introuvable - revient à nier toute relation à la fonction. En effet, une fonction se révèle toujours efficace, dès sa première apparition: le concept de roue a certainement commencé à exister mais pour une fonction précise à un moment donné. Ce concept a évolué depuis. Dans une classification naturelle on considère que la fonction est le point d'entrée dans l'histoire de l'évolution d'une structure, alors que cladisme nie cette relation, et remplace la notion de fonction (d'où dérive le terme d'homologie classique) par une notion relativiste d'homologie de filiation (voir ces mots ci-dessous). De la même façon - et sans quitter le sujet, car le cladisme compare souvent des séquences d'ADN qu'il qualifie de gènes - on pourrait dire qu'un gène n'a de sens que dans une cellule, à un instant donné de son histoire. La présence ou l'absence de la séquence d'ADN qui lui correspond sont insuffisants pour déduire autre chose qu'une possibilité de fonction: le gène est une unité fonctionnelle comme la génétique moderne ne cesse de le prouver (voir un des derniers commentaires en date sur la page d'accueil). Un même gène peut avoir plusieurs fonctions différentes dans divers environnements cellulaires. |
||||
|
|
|
homologie cladiste L'homologie de position (primaire) s'affirme de deux structures ayant la même organisation et les mêmes connexions avec les organes voisins (position anatomique). L'homologie de filiation (secondaire) s'affirme de deux structures similaires héritées d'un ancêtre commun (donc cette homologie est déduite a posteriori de l'arbre le plus parcimonieux). |
|
Deux structures homologues occupent une même position anatomique et ont une origine embryonnaire commune. Voir ancien cours de TS et nouveau cours de TS (en jachère), pour une discussion de ce concept. |
||||
|
|
|
|
||||||
|
2. La logique cladiste et les phylogénies |
|
|
||||||
|
|
|
2.1 Logique de la méthode pour classer des êtres vivants selon l'évolution |
|
|
||||
|
Les principes sur lesquelles repose la vérité de la classification évolutive cladiste du vivant Cette partie présente certains principes qui, je crois, sont acceptés par tous, d'autres, qui sont davantage sujet à discussion, et d'autres enfin qui sont des interprétations personnelles que j'essaie de préciser dans le paragraphe suivant. |
|
* Les êtres vivants ont une unité profonde qui s'exprime au niveau de leurs caractères qui se ressemblent (plus ou moins). |
|
Ce principe est le fondement de toute classification. Il n'est contesté par personne. Ce qui donne l'unité aux êtres vivants c'est le phénomène vivant, la vie, quelle que soit l'approche (philosophique) que l'on en fait. Il est clair que les similitudes matérielles (comme les molécules) n'expriment qu'une très faible part de cette unité. |
||||
|
* Les espèces évoluent (se transforment au cours des temps géologiques), ce qui doit aussi s'exprimer au niveau des caractères (des individus qui appartiennent à ces espèces) dont la ressemblance peut résulter de l'évolution. Aucune théorie particulière n'est supposée, ce qui est la grande force du cladisme. |
|
L'évolution est utilisée pour dire si un caractère dérive logiquement d'un autre ou non. Si l'évolution est un phénomène qui touche l'espèce son expression au niveau d'un caractère d'un individu n'est qu'une hypothèse théorique NON EXPÉRIMENTALE. |
||||||
|
|
|
Il est symptomatique que certains refusent le critère de ressemblance comme élément caractérisant les individus appartenant à une même espèce mais utilisent la ressemblance pour comparer phylogénétiquement des individus (voir espèce). |
||||||
|
* La construction de cladogrammes repose sur l'optimisation des relations logiques entre caractères (et donc des relations phylétiques entre individus). |
|
Optimiser c'est choisir le mieux (il y a une référence à un bien, donc à une finalité : il faut ordonner les possibles par rapport à une fin). Dans l'espace, pour aller d'un point à un autre, c'est le chemin le plus court (mais il faut se méfier de la représentation du temps comme un paramètre). |
||||||
|
* Les relations phylétiques mises en évidence entre les espèces ou les individus grâce aux caractères sont toujours relatives (aux choix du classificateur), ce qui est une faiblesse. |
|
Ce
sont des
relations
logiques
complexes de
similitude. |
||||||
|
* Le cladisme recèle une autre faiblesse: il ne tient pas compte d'une théorie de la variation. Le cladisme pourrait peut-être être complété en ce sens. |
|
En effet, le cladiste classe de façon optimale (vis-à-vis d'une relation logique évolutive de type filiation: B venant de A ou non) mais les mécanismes responsables de la variation lui échappent TOTALEMENT (il ne peut pas dire que A va évoluer en B ou C). C'est en quelquesorte une classification a posteriori puisqu'elle découle d'une filiation posée a priori. |
||||||
|
|
|
|||||||
|
analyse |
|
|
||||||
|
|
Le cladisme relie logiquement des caractères. |
|
Je rappelle que l'évolution porte sur les espèces et non les individus et encore moins les caractères (voir Qu'est-ce que l'évolution ? et la fiche Nommer-classer) |
|||||
|
|
|
|
|
1ère étape: le choix des caractères. |
|
|||
|
La méthode de classification cladiste est fondamentalement une comparaison... |
|
Le choix du terme caractère renvoie ici à un élément classificatoire et non héréditaire. En fait il s'agit de ressemblances (ou de plus ou moins grandes différences concernant des éléments ressemblants donc d'analogie au sens large). Les caractères pourraient très bien être appelés ressemblances ou similitudes. |
|
|||||
|
... entre caractères comparables, phylogénétiquement, c'est-à-dire homologues = portés par 2 individus ayant un ancêtre commun exclusif. On passe donc des caractères aux individus. |
|
Mais, pour le cladiste, tous les caractères ressemblants ne sont pas utilisables, il faut qu'ils traduisent des homologies "objectives". Les caractères choisis pour construire la phylogénie sont tous mis sur un plan d'égalité classificatoire. L'homologie de filiation (ou secondaire) est seule retenue comme objective et déterminante. Elle est définie par une relation logique dite de filiation (parenté exclusive) qui est une variante d'un principe d'unité des êtres vivants mais qui comporte une composante évolutive (phylogénétique), sans référence à une théorie particulière: deux caractères sont homologues s'ils sont portés par deux individus ayant une ancêtre commun exclusif (ce qui est déjà une expression du critère extra-groupe). Cette homologie NE pouvant être déduite des caractères eux-mêmes, elle est RELATIVE au choix des clades et s'énonce en fin d'opération classificatoire. Mais elle ne pourrait être affirmée si on ne disposait que d'un seul caractère. |
|
Homoplasies et homogénies sont des «concepts formalisés en 1870 par le zoologiste britannique Edwin Ray Lankester qui décomposa la notion d'homologie. Les homologies dues à la parenté sont appelées « homogénies » ; les autres, présentes chez diverses espèces sans pour autant avoir été héritées d'un ancêtre commun, sont appelées « homoplasies ». Depuis, on a restreint le sens de l'homologie à celui de l'homogénie, ce dernier terme tombant dans l'oubli.» Duris et Tassy (voir nommer-classer) |
||||
|
|
|
En
fait, cette
première
étape
est
précédée,
en plus du
choix de
l'individu (ou
du groupe :
ingroup)
que l'on
cherche
à
classer, par: |
||||||
|
Le clade regroupe des individus partageant des caractères homologues (et donc un ancêtre commun exclusif). |
|
Les individus regroupés par des caractères homologues forment un clade (nom masculin du grec klados = rameau): groupe monophylétique de référence. |
|
C'est ici qu'intervient une autre hypothèse qui ressemble toujours à la première mais qui est rarement énoncée: les caractères appartiennent à des êtres vivants et leur unité ou leur harmonie est l'expression de l'unité du vivant. Ainsi tous les caractères homologues choisis doivent permettre de construire UN arbre, qui est appelé l'arbre le plus parcimonieux en raison des hypothèses logiques qui seront présentées ensuite. |
||||
|
|
|
L'optimisation des relations logiques de l'arbre est une condition méthodologique qui prend deux orientations différentes selon la philosophie de son utilisateur: |
|
* pour la plupart des gens cette optimisation signifie que l'évolution, comme processus de filiation, est l'expression d'une relation des parties (individus) au tout (l'ensemble des êtres vivants ayant existé) |
|
* pour un réaliste, cette optimisation signifie que l'évolution comporte une finalité interne. |
||
|
|
|
Elle est d'autant plus acceptable qu'elle est impersonnelle et calculable. Cette optimisation est sous-entendue comme une caractéristique du vivant dans tous les processus naturels habituellement décrits au moyen de l'outil mathématique. Le cladisme n'y échappe pas. |
|
Cette optimisation se rapporte à l'évolution. On peut y voir une expression de la sélection naturelle revisitée (ce n'est plus un hasard mais bien une loi). On peut aussi davantage se retrouver dans le terme de finalité ou téléologie. Dans le cas où une caractère choisi ne serait pas homologue au sein du clade, on le verra apparaître dans deux clades différents, ce qui est incompatible avec l'hypothèse de départ (celle du clade ou celle de l'homologie, au choix, car il s'agit en fait de la même hypothèse). Les homoplasies, convergences, réversions ... et autres appellations des "non-homologies" sont ainsi dépistées. |
||||
|
|
|
|
|
2ème étape: La polarisation des caractères |
|
|
||
|
|
|
|
Lors de la polarisation des caractères homologues le cladiste décide si le caractère est primitif (valeur 0), c'est-à-dire hérité sans modification au sein d'un clade (et nul part ailleurs), ou si le caractère est dérivé (valeur 1), c'est-à-dire hérité avec transformation une seule fois entre deux membres du clade (et nul part ailleurs). |
|||||
|
Cette polarisation repose donc sur plusieurs hypothèses évolutives qui varient selon les caractères car ils sont soumis à deux critères: |
|
- le critère extra-groupe : |
|
- et le critère ontogénique : |
||||
|
|
|
|
il repose sur une hypothèse : plus un caractère apparaît tard dans le développement ontogénétique d'un organisme actuel, plus il est spécialisé et dérivé (apomorphe); plus il apparaît précocement, plus il est primitif (plésiomorphe). |
|
Remarque: |
|||
|
|
|
|
||||||
|
Remarque IMPORTANTE |
|
Il n'y a pas de temps pour le cladiste. Trois termes sont ambigus: |
|
|
||||
|
|
|
- le terme d'ancêtre ne désigne pas un fossile mais un concept (ce n'est pas un individu) qui regroupe un certain nombre de caractères. Quand on parle d'ancêtre "antérieur", il faut comprendre "plus éloigné en terme de similitude". Un ancêtre commun à deux groupes-frères est le point de rencontre logique le plus proche de tous les caractères propres à chaque groupe. Ce que le paléontologue interprète comme un indication bien sûr de l'emplacement du véritable ancêtre, historique, d'une lignée qui posséderaient les caractères portés par le clade. |
|
- les termes de primitif et dérivé ne signifient pas une succession temporelle. Seuls, les individus se succèdent dans le temps (hypothèse évolutive). Mais les caractères sont hiérarchisés non pas dans une succession temporelle (d'abord l'écaille puis la plume) mais dans une succession logique (le caractère plume de l'Archéoptéryx est éloigné logiquement de l'écaille de la Tortue mais est proche logiquement de l'écaille d'un reptile thécodonte). Le vocabulaire cladiste exact synapomorphie (partage de l'état apomorphe (du grec apo = loin de et de morphos = la forme), caractères éloignés) et plésiomorphie (du grec plésios = voisin et morphos = la forme, caractères voisins) évacue cette idée de temps avec justesse. Bien sûr, tout le monde va dire que la succession logique exprime la succession temporelle mais ce n'est pas exact. La succession logique exprime la plus parcimonieuse parenté, c'est-à-dire la phylogénie qui satisfait le mieux aux hypothèses logiques choisies. Si celles-ci ont une réalité, la phylogénie cladiste s'approche alors du réel, si elles ne sont qu'approchées, elle s'en éloigne. C'est en cela aussi que le cladisme est relativiste car il s'oppose à l'idée d'une connaissance certaine, non relative, par exemple historiquement définie. |
||||
|
|
|
N'est-ce pas jouer sur les mots ? |
|
NON, le cladisme est utilisé par le paléontologue qui accepte les relations logiques de similitudes comme raisonnables et sûres. Mais le cladisme se garde de porter le rôle de faiseur de vérité historique. Il s'arrête à ce qui est raisonnable logiquement, avec ses postulats. Alors que le raisonnement historique est autre: l'histoire suppose que les événements ont une lieu une seule fois, d'une certaine manière, même si cette connaissance est difficile à atteindre, elle repose sur une certitude... historique. |
||||
|
|
|
|
||||||
|
|
|
Dans
cette
étape
il y a un
basculement
des
caractères
aux individus.
Chaque
individu est
défini
comme un
ensemble de
caractères
et le graphe
relie les
individus (et
non plus les
caractères
qui ont
été
choisis,
comparés
et
polarisés). |
|
3ème étape : calcul du graphe Le cladogramme représente des liens de similitude (logiques) entre individus (seuls existants pour un nominaliste) qui ont (probablement) existé (et dont l'existence est attestée par les fossiles) ou qui existent (actuellement). |
|
Il est hors de question de détailler ici les règles logiques utilisées pour la construction d'un cladogramme : règles de parcimonie, UPGMA... Je n'en ai pas la compétence, ni le goût. |
||
|
|
|
Il semblerait, pour ce qui nous intéresse, qu'elles reposent tous sur le même principe logique exposé plus haut d'une optimisation du graphe obtenu à partir des caractères choisis. Cette optimisation est, encore une fois, selon la philosophie de l'utilisateur, l'expression d'une relation logique de l'évolution ou une finalité interne. |
|
|
||||
|
Remarque : |
|
On en arrive à un point délicat, que je présente ici sous une forme non définitive. |
|
Je crois comprendre différemment maintenant les mots de Mme Chandebois. Ce qu'elle stigmatise ici est peut-être ce que j'ai compris (grâce au travail d'André Pichot) comme étant une confusion entre évolution (phylogénie, relation de parenté) et histoire. Une fois encore, il n'y a pas de temps pour le cladisme donc pas d'ordre temporel mais uniquement logique. Étant
donnée
la profondeur
de
pensée
de Mme
Chandebois il
est peu
probable
qu'elle ai mal
compris le
cladisme. Sa
remarque
touche
à mon
avis un
problème
de fond du
cladisme qui
ne
s'intéresse
qu'aux
relations de
similitude
(le cladisme
est sans aucun
doute une
classification
qui
répartit
en classes
d'équivalence
des individus
en fonction de
leur
degré
de similitude)
et qui ne
suppose donc
seulement
qu'une
théorie
de la
similitude.
Celle-ci est
fournie d'une
part par l'unité
du vivant et
d'autre part
par une
théorie
de
l'évolution.
Mais cette
théorie
évolutive
nécessaire
au cladiste ne
s'intéresse
qu'à la
parenté
et
évacue
le second
aspect de
l'évolution
qui est celui
de la
variation.
Certes, la
filiation est
définie
comme
transformation
dans le cas
d'apomorphie,
mais celle-ci
me
paraît
incapable de
décrire
la
spéciation.
Car pour
comparer en
termes de
similitude il
faut deux
membres, or il
n'y a rien,
dans la
classification,
au delà
de
l'espèce.
C'est pour
cela que le
cladisme
échoue
aussi pour les
premières
étapes
du vivant. Il
ne peut
comparer
l'hypothétique
première
cellule avec
des
éléments
non
organisés
et non
vivants. |
||||
|
|
|
R. Chandebois: «D'après les "arbres logiques" les embranchements auraient été créés d'abord. leur genèse achevée, celle des classes aurait commencé, puis celle des ordres aurait emboîté le pas, et ainsi de suite jusqu'à l'individualisation des espèces qui, pour cette raison, se trouvent toutes à la périphérie de l'arbre. (...) La filiation entre les embranchements, les classes, les ordres, n'a pu se faire autrement que par des espèces particulières. C'est sur ce point que la schématisation cladiste est défectueuse. Sur le dessin elle est fausse, dans le texte elle est ambiguë » (Pour en finir avec le darwinisme; une nouvelle logique du vivant, Éditions espace 34, 1993, p 111-112). |
|
|||||
|
|
|
Une fois établie la relation à l'ordre, à la classe, à l'embranchement, l'individu reste plus que la somme des similitudes. Pour que le cladisme puisse produire des phylogrammes qui tiennent compte à la fois de la stabilité et de la variabilité dans l'évolution il faudrait une représentation de l'individu en tant que tel et non pas comme "le phénomène de ses relations avec les autres". Est-ce possible sans un réalisme ? Je laisse ce travail en suspens... Mais le cladisme sera sans aucun doute dépassé. |
||||||
|
Un cladogramme n'est pas une phylogénie |
|
La difficulté est de comprendre la liaison du graphe avec l'histoire évolutive ou phylogénie au sens strict que, comme paléontologue, nous souhaiterions établir. Si l'on place verticalement le temps le long du graphe on remplace la succession logique par une succession temporelle, ce qui est parfois acceptable mais souvent l'est moins. En tout cas le sens d'un tel passage n'est jamais donné par le cladisme, il faut lui trouver une justification extérieure. |
|
Le cladogramme nous présente des individus, seuls réels, qui sont situés aux extrémités des branches (liens de similitude les plus forts) ou au voisinage des nœuds (points de divergence ou plutôt convergence de groupes de similitudes) représentent des innovations qui peuvent permettre de nommer les clades qui en partent...; il n'y a pas de proportionnalité entre la distance entre deux individus et leur degré de parenté (proximité phylogénique) qui est seule représentée par les droites et les nœuds. Même si certaines présentations scolaires , à mon avis confuses, s'efforcent d'ajouter cette dimension, que je crois savoir extérieure au cladisme. |
||||
|
Les arbres ne sont pas comparables entre eux |
|
Pour essayer d'y voir clair il faut bien garder présent à l'esprit la valeur relative de l'attribut évolutif affecté à chaque caractère (primitif-évolué). Cette valeur n'a de sens que dans une phylogénie posée comme point de départ (provisoire). De plus la valeur change pour un même caractère selon les groupes-frères considérés. Cette valeur relative à un arbre de chaque caractère, fait que les arbres ne sont pas comparables entre eux. Lorsque l'on construit un cladogramme, celui-ci n'est qu'une représentation de l'hypothèse de départ *, illustrée par les fossiles étudiés, ou plutôt par les caractères étudiés dans les groupes étudiés. Un cladogramme n'apporte AUCUNE PREUVE de la filiation entre des organismes fossiles ; il n'est en est que l'illustration. * pour essayer de montrer ce point important je préciserai cette hypothèse dans les graphes analogiques pris en exemples ci-dessous |
||||||
|
Par exemple comparez l'intérêt pédagogique des deux classifications des animaux très simplifiées données dans mon cours de seconde: une classification cladiste est à mon avis plus difficile à mémoriser et moins signifiante qu'une classification phylogénétique "embryologique" classique. |
|
Certes cette méthode présente l'avantage de rendre accessible à tous la classification; désormais, n'importe quel étudiant peut à l'aide d'un logiciel, et s'il dispose de la matrice actuellement acceptée par les sommités cladistes, réaliser une classification cladiste qui aura la même valeur que celle du plus grand spécialiste. Il y a cependant une double tromperie: |
|
* la matrice, seule à refléter les hypothèses évolutives, échappe à l'étudiant et donc c'est toute idée de travail scientifique qui lui échappe. * lors de la polarisation des caractères, je serais tenté de dire - de façon peut-être un peu excessive - que les hypothèses hardies d'ordre paléontologique ou embryonnaire, ou même d'ordre moléculaire, sont remplacées par la règle la plus aliénante qui soit: il n'y a pas de vérité. C'est ce côté relativiste de la méthode cladistique qui me paraît le plus dangereux. On pourrait dire que la cladogenèse remplace la recherche d'une phylogénie universelle par une appréciation phylétique relative à un ensemble de caractères donné dans une certain nombre de groupes donnés. Il y a une forte tendance à relativiser la valeur de la phylogénie, tout en affirmant que cette méthode est la seule scientifique et la seule réfutable, puisqu'elle change en fonction des caractères et des groupes considérés : réfutable ne veut pas dire provisoire ou encore moins relative. Je crois que l'on peut dire que, à plus d'un titre, le cladisme est une phylogénie relativiste. |
||||
|
|
|
|
||||||
|
|
|
2.2 Discussion |
|
|
|
|
||
|
|
|
Les premières questions soulevées ci-dessous appartiennent à la première version de cette page. Je suis conscient qu'elles n'intéressent plus grand monde. Maintenant tous les biologistes utilisent les classifications cladistes et ne se posent plus de questions. Ils se sont appropriés l'outil et les emplois nouveaux que l'on en fait semblent plus intéressants (voir prochaine partie: recherche) que les questions que l'on peut se poser sur la fiabilité de l'outil. Cependant, je persiste à croire que les actuelles phylogénies ne sont ni fidèles au cladisme (qui est anhistorique) ni cohérentes avec la recherche d'une histoire du vivant (voir nommer-classer). |
||||||
|
parti-pris
|
|
Que l'on pardonne parfois des affirmations trop dogmatiques: je suis bien conscient, en tant que pédagogue, que la faute m'en revient à moi et non aux scientifiques, qui, d'ordinaire, n'érigent pas leur théorie en dogme. Comme le dit si justement G. Canguilhem (dans la théorie cellulaire) en se référant au fixisme: «...pourquoi l'historien des sciences présenterait-il Linné comme le responsable d'une rigidité doctrinale qui incombe à la pédagogie plus qu'à la constitution de la théorie ? ». En caricaturant certaines théories scientifiques, en les dogmatisant, je sais que je ne leur rend pas justice. |
|
Mes critiques - basées sur mes sources, citées, scolaires pour certaines - ne visent peut-être que certaines idées qui ne reflètent pas tous les courants cladistes. Je serais ravi d'avoir d'autres ouvertures |
||||
|
naïveté
|
|
Le
cladisme,
présenté
par ses
actuels
promoteurs, n'est
pas qu'un
outil de
présentation
de phylogénies. L'obstacle majeur est celui du temps car la phylogénie cladiste est une représentation de relations logiques et non pas un arbre généalogique avec le temps qui occuperait l'une des dimensions du plan de représentation de l'arbre. |
|
L'ambition
(démesurée)
des cladistes
est de fournir
LA
MÉTHODE
(unique) de
classification
MODERNE et
ÉVOLUTIVE
de TOUS les
êtres
vivants. |
||||
|
commodité |
|
Les classifications cladistes ne sont pas les seules classifications qui tiennent compte des caractères moléculaires. |
|
Ceux-ci peuvent entrer dans TOUTES les classifications, mais il faut pour cela expliciter des hypothèses qui sont souvent implicites pour les classifications cladistes. Par contre les comparaisons entre molécules ont surtout été réalisées avec l'outil cladiste qui a été mis au point avec les données obtenues par ces études moléculaires. Le cladisme reste donc un outil tout à fait adapté pour des études comparatives de séquences. D'ailleurs la considération des étapes de la méthode cladistique à partir de séquences moléculaires permet pédagogiquement de mettre en évidence les limites de la méthode. - Il est clair qu'il n'y a pas de temps : ni expérimental (une séquence ne dérive d'une autre que par pure hypothèse évolutive, jamais grâce au résultat d'une expérience) ni historique (les séquences ne sont pas datées, sauf quelques rares exemples de molécules anciennes...). - Il est clair que les relations logiques établies entre séquences vont dépendre des mécanismes "évolutifs" (il ne s'agit que d'évolution moléculaire... une théorie très limitée, bien incapable de rendre compte de l'évolution des espèces) supposés, pris comme hypothèse et inclus dans le diagramme (séquence stable, taux de mutation (horloge moléculaire), duplication de gènes, transposition...), mais en aucun cas commentés (pour la communauté, comme on peut le faire en systématique évolutive) ou validés (c'est à mon avis un point majeur: le cladogramme illustre mais ne prouve rien). - Il est clair que les règles de construction de l'arbre vont reposer sur des lois logiques de minimisation des différences qui n'ont rien de biologique ou d'évolutif... |
||||
|
évolutionnisme |
|
Les classifications cladistes ne sont pas les seules classifications évolutives. |
|
Toute
classification
qui
intègre
des
données
paléontologiques
peut se dire
évolutive,
mais elle ne
l'est vraiment
que si elle
repose sur une
théorie
évolutive.
Le cladisme en
suppose une
mais ne
l'utilise que
dans la
polarisation
de ses
caractères,
elle est donc
souvent
cachée. |
||||
|
|
|
|
|
|
|
|||
|
|
Les classifications cladistes sont évolutives mais ne sont pas toutes darwiniennes... |
|
... dans le sens où le cladisme reposerait sur une théorie darwinienne de l'évolution car ce n'est qu'une méthode qui suppose une évolution , quelqu'en soit le déterminisme, s'il existe. Par sa représentation des nœuds correspondant à des innovations biologiques et par son refus de la recherche d'ancêtres, le cladisme pose au contraire des difficultés à une interprétation anagénétique darwinienne (voir ces mots dans l'ancien cours de terminale (§4b): l'anagénèse, progressive et continue, s'opposant à la cladogénèse, discontinue ; cette distinction n'est d'ailleurs peut-être pas aussi moribonde qu'on veut bien le dire, même si l'évolution est maintenant surtout vue comme buissonnante). |
|||||
|
relativisme
? |
|
Du point de vue philosophique, la méthode cladistique qui nous est présentée actuellement est héritière du nominalisme et, à mon avis, fréquemment adossée au relativisme philosophique, comme je vais essayer de l'établir. Je crois que, si l'on ne peut pas s'affranchir de la relativité des liens logiques entre les éléments d'une classification cladiste , on reste en droit, par contre, de ne pas accepter le relativisme philosophique qui va souvent avec. |
|
Contrairement
à ce
que certains
cladistes,
philosophiquement
matérialistes,
affirment,
l'idée
d'une
classification
naturelle,
naturellement
ordonnée,
n'est pas une
idée du
passée,
une folie
d'essentialistes
bornés,
mais le
fondement de
toute
classification
sensée.
Et cela
n'empêche
pas d'utiliser
la cladistique
comme une
méthode
scientifiquement
intéressante
(de par les
liens qu'elle
établit
dans la
communauté
scientifique). |
||||
|
|
|
L'ambiguïté est peut-être entretenue par les promoteurs-vulgarisateurs des classifications cladistes qui rejettent fortement l'essentialisme sans pour autant adhérer philosophiquement à un relativisme anti-réaliste. Leur position plus ou moins scientiste est venue ainsi ternir l'image d'une cladistique moins marquée philosophiquement par le relativisme, et qui reste, à mon avis, possible. (voir citation de R. Blanchette ci-contre) |
|
« De fait, incapables de sortir du dilemme que leur posent leur acceptation de la condamnation de l'essentialisme et leur hésitation à embrasser pleinement l'anti-réalisme épistémologique, les écoles [de classification] se contentent de proposer des approches de la prise de décision différentes en reportant leur attention sur la question de l'objectivité en science ». (thèse de Richard Blanchette, Le problème de la classification en zoologie, Université Laval (Québec), PhD, juin 2002) que l'on peut trouver in extenso au format PDF sur internet à l'adresse: http://www.theses.ulaval.ca /2002/20168/ 20168.pdf) Partie
précédent
la citation
ci-dessus, qui
ouvre le
débat
sur les autres
classifications: |
||||
|
|
|
Remarques : |
|
|
|
|
||
|
|
|
J'ai trouvé dans mes sources deux références au nominalisme: Bruno Chanet qui affirme que le principe de parcimonie est hérité de Guillaume d'Occam (Bruno Chanet, Biologie-Géologie n°3-1997, p 507) et Guillaume Lecointre et Hervé le Guyader qui dans leur introduction, affirment que «la plupart des phylogénéticiens ont, à l'égard du vivant, une attitude nominaliste qui fut celle de Darwin, aux antipodes de l'essentialisme» (Classification phylogénétique du vivant, Guillaume Lecointre et Hervé Le Guyader, Belin, 2001,p 18). |
|
Les auteurs de Classification phylogénétique du vivants considèrent que «croire que l'évolution a un but , a , pour corollaire, de croire que la nature intime des êtres, l'essence, d'un point de vue philosophique, précède l'existence » (p 17). Derrière cette phrase caricaturale je pense que les auteurs se réfèrent à Popper en utilisant le terme d'essentialisme: (Popper, Conjectures et Réfutations, chap. 3,3). Ce dernier utilise ce terme pour qualifier (par opposition à l'instrumentalisme et à la définition popperienne d'une démarche par conjecture et réfutation) que l'objet de la connaissance, même scientifique, est l'essence des choses, cachée derrière les apparences; Dans le cadre d'une acception popperienne, l'essentialisme a 3 conséquences que l'on peut résumer ainsi: toute science possède un objet fixe, il existe une vérité, sens et vérité sont synonymes. |
|
Cependant,
l'acception
popperienne du
mot
essentialisme
n'est pas
l'acception
première
de philosophie
opposée
à
l'existentialisme
sartrien.
Quoiqu'il
existe aussi
une
utilisation
plus subtile,
en logique,
pour
désigner
une assertion
selon laquelle
s'il y a des
lois vraies
(logiquement),
elles
décrivent
non seulement
ce qui est
(en dehors
de la
façon
dont nous les
percevons)
mais aussi ce
qui doit
être
(propriétés
nécessaires
ou
essentielles);
mais ces
vérités
nécessaires
ne sont pas
connaissables
a priori mais
a posteriori
pour un
essentialiste.
La
définition
qui en est
donnée
(l'essence
précède
l'existence)
ne vaut que
dans un
discours
logique. On
pourrait dire
que Platon est
essentialiste,
mais pas
Aristote, ni
moins encore
Saint Thomas. |
||
|
|
|
Le seul article, disponible sur internet, et qui s'élève un peu, au sujet de l'ouvrage de Lecointre-Le Guyader est de Claude Lévi-Strauss (Guillaume Lecointe & Hervé Le Guyader, Classification phylogénétique du vivant. Illustrations de Dominique Visset. Publié avec le concours du Centre national du livre. Paris, Belin, 2001, 543 p., annexes, bibl., index, tabl., L'Homme, 162, Questions de monnaie, 2002 à l'adresse: http://lhomme.revues.org/ document169.html |
|
|
||||
|
|
|
|
|
|
||||
|
|
|
Si le cladisme "pur" (anhistorique) est mort on peut identifier au moins deux prolongements (souvent mélangés) : le cladisme "statistique" et le cladisme "historique". |
||||||
|
|
|
3.1 cladisme et statistiques |
|
|
||||
|
Le seul qui choisit c'est le chercheur... pas la machine
Comprendre et utiliser les statistiques dans les sciences de la vie, B. Falissard, Masson, 2005 Un livre qui pose de bonnes questions dans certains domaines (voir site auteur) mais presque sans mathématiques, ce qui en décevra certains |
|
Si
les
statistiques
se retrouvent
à tous
les niveaux de
l'analyse
cladistique,
ce n'est que
récemment
(au
XXIème)
qu'elles ont
pris une telle
ampleur et il
y a un risque
d'erreur car
rares sont les
biologistes
à
maîtriser
l'outil aussi
bien qu'un
spécialiste
(mais est-ce
souhaitable
?). |
|
* Le fait de faire tourner un programme qui "choisit" l'arbre optimal pour vous (il calcule celui qui possède les caractéristiques que VOUS avez choisies) n'est pas sans induire une certaine déconnexion entre le résultat (qui est toujours comparé avec ce qui était attendu) et l'ensemble des hypothèses (parfois bien cachées) nécessaires à l'obtention de celui-ci. |
|
* Les statistiques sont par contre employées avec de plus en plus de pertinence pour construire des caractères simples discontinus (morphologiques par exemple..., voir exemple) à partir de regroupements de plusieurs caractères complexes dont certains peuvent être continus, ou encore pour extrapoler des caractères manquants dans certains groupes... Cette fois ces méthodes statistiques de construction sont assez classiques et ne demandent pas un niveau de connaissance mathématiques trop élevé pour un naturaliste. |
||
|
|
|
|
|
|
|
|
||
|
|
|
3.2 cladisme et phylogénie |
|
|
||||
|
cladisme et typologie |
|
Au sens premier les taxons des cladogrammes doivent être des individus et non des espèces, concept non expérimental. Ainsi chaque taxon se trouve ainsi réduit à être représenté par une somme de caractères. Ne pourrait-on voir ici une certaine résurgence de l'espèce typologique, tant décriée par certains ? |
|
évolution des caractères |
|
L'évolution
touche les
espèces
et non les
individus.
Encore moins
les caractères.
Les
caractères
des individus
peuvent-ils
être
interprétés
phylogénétiquement
? |
||
|
représenter le temps et même la vitesse d'évolution |
|
Dans
la
représentation
cladistique
classique de
l'évolution,
les traits
reliant les
taxons
représentent
une relation
logique de
ressemblance
(similitude)
évaluée
statistiquement
(caractères
polarisés
identiques les
plus
nombreux...).
La
ressemblance
se traduit par
une distance
proportionnelle
à cette
ressemblance
sans qu'il
soit possible
d'invoquer un
mécanisme
de changement
brusque (de
type catastrophique
au sens de René
Thom). Les phylogrammes moléculaires qui sous-tendent des mécanismes génétiques moléculaires évolutifs (mutations, duplications...) utilisent clairement le temps et proposent même la notion d'horloge moléculaire. Ces représentations ne peuvent clairement plus être considérées comme des cladogrammes, même si des analyses de type cladistique sont réalisées sur les séquences. |
|
L'utilisation de datations, avec des fossiles par exemple, conduit aussi à des phylogrammes qui ne sont plus en accord avec les principes des cladogrammes (rotation possible autour des nœuds par exemple). |
|
Mais il existe d'autres représentations qui contiennent le temps à partir d'un vrai mécanisme historique évolutif, mais non limité aux molécules. C'est la voie proposée par André Pichot avec sa divergence évolutive mesurant une vitesse d'évolution. 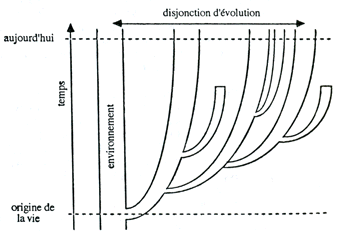 Histoire de la notion de vie, André Pichot |
||
|
|
|
|
|
|
||||
|
hérédité et évolution page spéciale à partir du travail d'André Pichot: Hérédité, reproduction et évolution |
|
Quels sont les liens entre l'hérédité, la reproduction, la descendance et l'évolution ? Depuis Weismann il existe une vraie confusion entre généalogie et phylogénie, entre hérédité et mémoire exprimée notamment chez Haeckel (avec les mots d'André Pichot : «... l'hérédité est supposée être une sorte de mémoire ayant gardé la trace de la phylogénèse et, en commandant le déroulement de l'ontogénèse, elle fait de celle-ci une récapitulation de la phylogénèse (voir aussi page annexe)». Il est très difficile de sortir de sa tête cette séparation entre le matériel génétique héréditaire (associé traditionnellement aux chromosomes mais que l'on tend à élargir aujourd'hui aux ARN, voire à certaines protéines...) et processus acquis (souvent nommés à tort épigénétiques) stockés on ne sait où... |
|
Quand à la reproduction elle a été bien trop longtemps enseignée comme un simple moyen de transmettre des chromosomes (quand ce ne sont pas les gènes...), ce qui occulte sa dimension proprement évolutive, du fait de son rattachement à l'espèce et non à l'individu. |
||||
|
|
|
|
|
|
|
En travaux |
||
|
4 . Des exemples de "vrais" et de "faux" cladogrammes |
|
|
||||||
|
Plan: http://tolweb.org/ : l'arbre de la vie... ou plutôt des arbres. Les arbres présentés mêlent à la fois les cladogrammes et le temps de façon plus ou moins claire. |
|
Remarques: La présentation d'un arbre phylogénétique (phylogenetic tree) relève donc de choix discutables (et discutés) mais cache parfois une erreur de compréhension de l'intérêt ou même des fondements du cladisme. Dans ce paragraphe je présenterais, au fur et à mesure, des cladogrammes-arbres pouvant être utiles.... |
||||||
|
|
|
|||||||
|
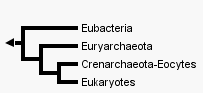
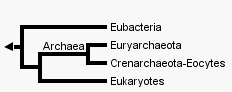
|
Deux arbres qui sont en fait des cladogrammes (on est bien incapable de placer le temps de façon certaine, même si l'on pense que les eucaryotes devraient apparaître après les procaryotes.... mais cela aussi se discute...). Deux
hypothèses
sur l'origine
des Eucaryotes
: les
Archées
(bactéries
"archaïques")
forment-elles
un clade ? |
|
Les
Eubactéries
comprennent
les
bactéries
au sens
classique
d'où
seraient
dérivées
les
mitochondries
et les
chloroplastes
des cellules
eucaryotes.
Un
très
intéressant
article
d'histoire des
sciences par Jan
Sapp (en
libre
accès)
qui replace
historiquement
la
signification
de la
séparation
procaryotes-eucaryotes: |
|||||
|
Comme il n'existe pas de définition consensuelle de l'espèce en bactériologie, il n'existe pas non plus de classification unique et consensuelle des procaryotes (voir par exemple le page de J.P. Euzéby). Les taxinomies phénétique, moléculaire ou encore chimique, sont encore très employées. |
|
|||||||
|
|
|
|||||||
|
|
|
Pour naviguer dans ce cladogramme pédagogique de T. Mike Keesey de 1999 mis en forme par Jeff Poling (qui insiste bien sur son côté provisoire et non consensuel), allez à la page d'accueil plus cliquez sur les nœuds rouges du cladogramme pour obtenir des parties détaillées des clades réunis au niveau des nœuds. Les
taxons "incertæ
sedis" (notés
i.s.) sont
des fossiles
plus ou moins
fragmentaires
inclassables.
On leur
attribue
parfois une
classification
propre
(parataxinomie)
plus ou moins
signifiante. |
|
Dinosauria Cladogram : http://www.dinosauria.com /dml/clado /ci.htm |
||||
|
|
|
|||||||
|
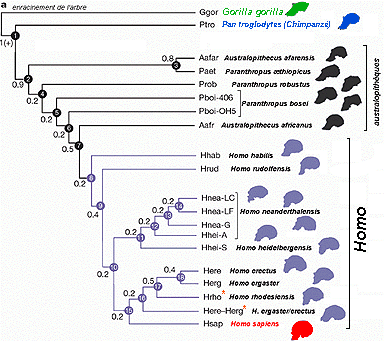
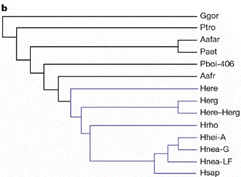
González-José, R., Escapa, I., Neves, W.A., Cúneo, R., Pucciarelli, H.M. (2008). Cladistic analysis of continuous modularized traits provides phylogenetic signals in Homo evolution. Nature DOI: 10.1038/nature06891 (version gratuite en ligne accessible le 6/06/2009)
cours de 1ère l'évolution de l'homme |
|
Un
autre exemple
très
pédagogique
(et attirant)
bien que fort
difficile est
celui de la
lignée
humaine. |
||||||
|
Tableau
1 1.
Aafa (A.L.
444-2)
(reconstruction)
Australopithecus
afarensis (
≥ 3.7 - 3.0
Ma)
|
|
|
||||||
|
|
|
|||||||
|
|
Le
principal
résultat
est que le
genre Homo
correspond
à un
clade
(nœud
8),
c'est-à-dire
un groupe
monophylétique
(avec un
ancêtre
commun). C'est
une des
constantes des
résultats
de la
paléoanthropologie.
L'état
ancestral des
caractères
pour le
nœud 8
est
représenté
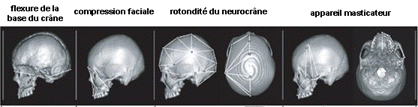
ci-dessus : on
observe une
flexure de la
base du
crâne
plus
prononcée,
une face plus
rétractée
et une
augmentation
de la
rotondité
neurocrânienne. |
|||||||
|
|
|
Data
acquisition
and geometric
morphometrics. Cladistic
analysis. |
|
Acquisition
des
données
et mesures
géo-morphométriques Analyse
cladistique |
||||
|
En travaux |
|
(1)
'analyse
en composantes
principales
(ACP) est une
technique
d'approximation
qui consiste
à
réduire
un grand
nombre de
variables
(sans
condition
particulière
sur celles-ci)
à un
ensemble de
variables
indépendantes
(les
composantes
principales)
calculées
comme
étant
les variables
qui
présentent
la variance
maximale. |
||||||
|
|
|
|
||||||
|
|
Le
but de cet
exemple est
double : Objectif clairement affiché: concilier les données paléontologiques (chronologie, anatomie...) avec les données moléculaires ayant donné lieu à des cladogrammes qui sont restés, pendant de nombreuses années, en franc désaccord avec les résultats des méthodes classiques. |
|
Les ancêtres des hippopotames, J-R. Boisserie et al. La Recherche, 450, mars 2011, pp 52-55 : encore un article vraiment pédagogique de La Recherche et accessible à des élèves de lycée. On notera que cladogramme et chronologie sont ENFIN séparées comme l'exige la méthode cladiste. |
|||||
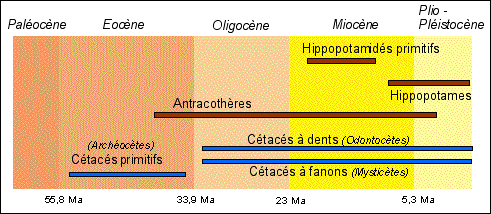
|
Ce sont les cladogrammes construits UNIQUEMENT à partir de caractères moléculaires qui ont été les déclencheurs d'une recherche toujours en cours pour arriver à un consensus sur l'origine des cétacés et partant des hippopotamidés. |
|
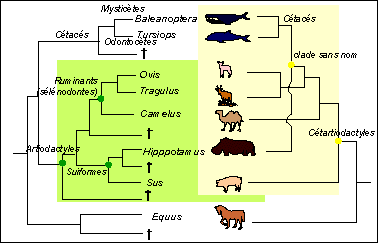
Avant 1990 les résultats obtenus à partir des quelques séquences d'aa comme la β-hémoglobine ou la ribonucléase indiquaient déjà un rapprochement entre les Artiodactyles et les Cétacés. Mais le gros du travail a été réalisé entre 1990 et 2001. Gatesy rapporte en 2001 que plus d'un million de paires de bases d'ADN provenant de cétacés ou d'espèces apparentées ont été collectées dans ces années (séquences de gènes de protéines ubiquistes, séquences de gènes mitochondriaux, nombreux autres gènes). Des méthodes complémentaires d'analyse de génome comme les hybridations ADN-ADN et l'insertion de transposons ont aussi été utilisées. Les cladogrammes obtenus à partir des données moléculaires allaient TOUS dans le sens de l'origine commune entre Cétacés et Artiodactyles. Un nouveau groupe - les Cétartiodactyles- était envisagé. |
|
|
||||
|
Comme les hippopotames étaient d'incontestables artiodactyles, cela fournissait une origine commune aux artiodactyles et aux cétacés. Mais ces données moléculaires étaient en désaccord avec les données paléontologiques, car aucun fossile ancien de cétacés (datés d'environ 53 millions d'années) ne présentait un élément anatomique toujours présent chez les artiodactyles: l'astragale (os de la rotule) à double poulie. Les mammifères terrestres artiodactyles sont un ordre dans lequel on classait les Hippopotames, les Suidés (cochons et sangliers) et de nombreux autres mammifères dont l'axe du membre passe entre les doigts III et IV comme les Ovins ou les Camélidés). Les Cétacés avaient une origine incertaine même si les fossiles, formant le groupe des Archéocètes, extraits de sédiments datés de l'Eocène-Miocène, montraient d'évidents rapports avec les Mammifères terrestres. On penchait même parfois pour une origine différente entre les Odontocètes (Cétacés à dents: dauphins, orques et autres planistes (dauphin de rivière)) et les Mysticètes (Cétacés à fanons comme les baleines).(in Grassé, Précis de zoologie, t3, Masson, 1977). |
|
|
||||||
|
En 2001, deux équipes « publièrent des analyses de deux squelettes quasi complets de cétacés primitifs datés de 50 à 45 Ma et possédant des caractéristiques d'artiodactyles : quadrupèdes, avec des pattes arrière parfaitement développées, [] et un astragale indéniablement pouvu de deux poulies.» Il ne restait plus qu'à relier les hippopotames aux Cétacés grâce aux fossiles. Ceci a été réalisé par une équipe française entre 1997 et 2005. En utilisant la méthode cladiste de comparaison de nombreux caractères anatomiques de fossiles de tous les groupes qu'ils souhaitaient relier, ils ont pu enraciner les hippopotames primitifs au sein du groupe fossile des Anthracothères. Pour parfaire leur analyse, ils intégrèrent les dents de nombreux hippopotamidés fossiles (notamment Kenyapotamus, le plus ancien fossile du groupe daté de 16 à 8 Ma) à leurs arbres et c'est en 2010 qu'ils établirent de façon rigoureuse que les hippopotames dériveraient de certains Anthrocothères, d'autres évoluant vers les Cétacés. Depuis 2001 environ on peut donc dire que les études anatomiques à partir des fossiles et les études moléculaires sur les espèces actuelles n'ont cessé d'aller dans le même sens. Les découvertes se succèdent et confortent les cladogrammes qui deviennent des phylogrammes intégrant les données actuelles et fossiles et le temps. |
|
Un
truc pour ne
plus se
tromper dans
l'orthographe
d'hippopotame: |
||||||
|
|
|
|
||||||