|
||||||
|
Sources
générales |
||||||
|
Plan |
|
|||||
|
En travaux 05/2010 |
||||||||
|
3.1
- Quelques points d'histoire autour de l'opéron
lactose
|
sources principales de ce
chapitre: |
|||||||
|
|
||||||||
|
L'étude, par François Jacob et Jacques Monod, chez Escherichia coli, et à partir de mutants, de ce qu'ils appelèrent "l'opéron lactose", groupe de deux gènes de structure très voisins et régulés ensemble (galactosidase et perméase, nécessitant notamment un gène-opérateur voisin), a été récompensée en 1965 par le Prix Nobel de physiologie et de médecine (avec André Lwoff «pour leurs découvertes sur la régulation génétique de la synthèse d'enzymes et de virus»). |
Le modèle présenté alors n'a cessé d'être perfectionné mais il est encore souvent présenté de façon très similaire à celle imaginée par ses premiers concepteurs. Ce modèle reste sans aucun doute paradigmatique (en ce sens qu'il ne nécessite plus d'explication puisque tout un chacun connaît les a priori moléculaires qui le fondent), mais doit être dépassé. Du point de vue historique on peut
accéder à certains des articles de Jacob,
Monod et d'autres membres plus ou moins oubliés de
leurs équipes. |
|||||||
|
La
physiologie bactérienne d'abord... |
|
|||||||
|
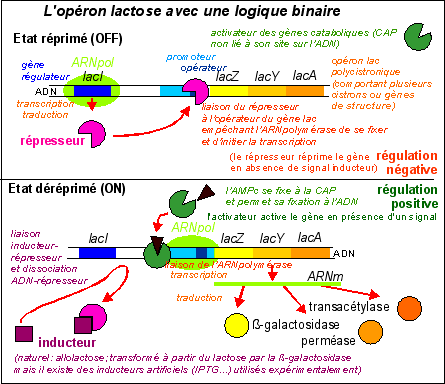
L'opéron
lactose comporte 3
gènes
(lacZ, laY et lacZ, qui forment un
cistron) codant pour 3
protéines nécessaires chez Escherichia
coli à l'utilisation du glucose comme source de
carbone |
|
Il est, de plus, déterministe ou instructionniste (si on utilise un vocabulaire plus lié à l'information génétique) dans la mesure où chaque susbtance chimique présente un rôle précis déterminé par sa concentration et sa localisation à un instant donné.
|
||||||
|
|
||||||||
|
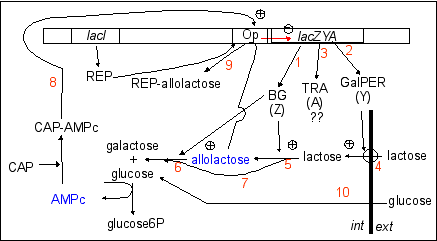
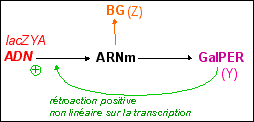
Un schéma des régulations autour de l'opéron lac présentant les principales voies métaboliques citées ci-dessous |
P.S. La protéine synthétisée par le gène de contrôle lacI (qui est aussi nommé "gène répresseur" ou simplement "répresseur"), est parfois appelée inducteur (IND). La protéine est appelée ici REP pour éviter des confusions avec les inducteurs métaboliques qui induisent les cellules à produire de la ßgalactosidase (qui sont les sucres comme le glucose ou le lactose ou encore l'allolactose ou le TMG... et se fixent à la protéine REP en la libérant de l'ADN). |
La ßgalactosidase (BG) est au
centre de la régulation avec l'allolactose qui non
seulement peut être repris par la Bgalactosidase pour
donner du glucose et du galactose (6), mais aussi se fixer
(9) au répresseur (REP) pour aller libérer
l'opérateur (Op) et permettre ainsi la transcription
du gène de la ßgalactosidase. |
||||||
|
Les expériences de M. Cohn et A. Novick réinterprétées par leurs auteurs (associés encuite à M. Weiner et K. Horibata respectivement)
Au sujet du rôle de l'Institut Pasteur dans l'émergence de la biologie moléculaire voir par exemple Georges N. Cohen (en anglais) http://www.jbc.org/ content/ 277/52/ 50215.full |
Fait courant en histoire des sciences, la
mise en place des premiers modèles physiologiques
montre la complexité et l'imbrication des
événements et des influences entre personnes.
Sans prétendre à une étude historique
j'ai cru comprendre que deux éléments majeurs
peuvent être retenus: |
Novick A., Weiner M., 1957. Enzyme induction as an all-or-none phenomenon. Proc. Natl. Acad. Scie. USA 43, 553-566 (accès libre) Inhibition
by glucose of the induced synthesis of the
ß-galactoside-enzyme system of Eschericchia coli.
Analysis of Maintenance, Melvin Cohn
and Kengo Horibata, 1959 J Bacteriol.
1959 November; 78(5): 601&endash;612.
article en libre accès
pour le premier article
|
||||||
|
maintenance
de phénotypes différents
dans des populations génétiquement
identiques Le phénomène de maintien (en anglais "maintenance") a été observé lorsque deux populations de même génotype, sont placés dans des conditions différentes et se maintiennent indéfiniement avec des phénotypes différents (une des populations produit de la ß-glalactosidase - elle est dite "induite" -, l'autre non). |
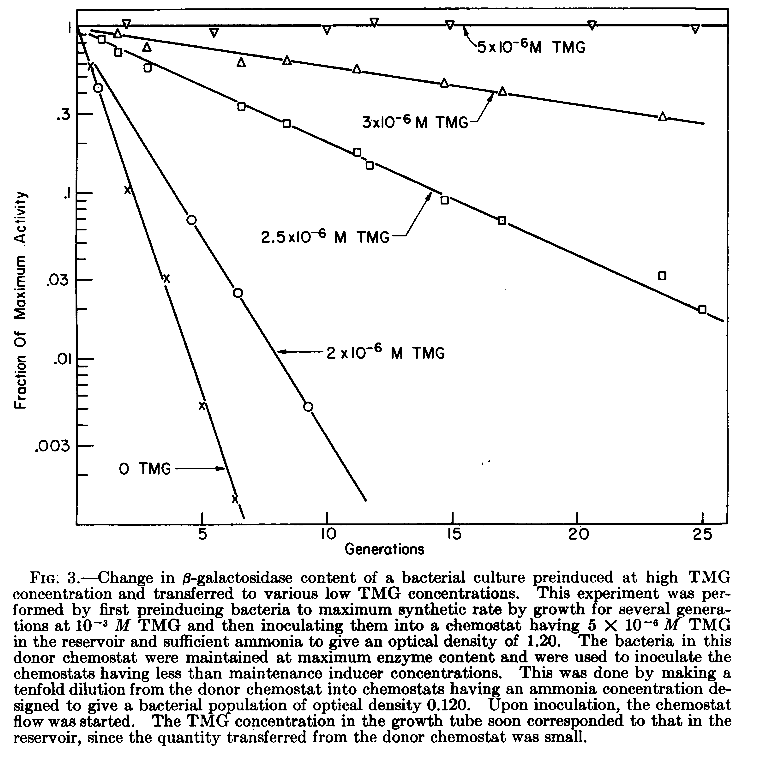
Dans les articles de Monod et d'autres
chercheurs ayant exploré le phénomène
d'induction par le TMG, l'adapation enzymatique d'une
population bactérienne sous l'influence d'un
inducteur gratuit est nommée
"pré-induction". Monod
expliquait ce phénomène par la
synthèse, progressive puis maximale (une fois un
seuil atteint), des deux enzymes: ßgalactosidase et
perméase. La quantité intracellulaire de TMG,
lorsque l'induction maximale était atteinte,
étant environ 100 plus élevée que dans
le milieu extérieur. |
Remarque: « Under normal
conditions glucose inhibits the induction of both
the enzyme and the permease. However, Melvin Cohn
discovered that if the bacteria are pre-induced by
TMG and glucose is then added, there is no
inhibition by glucose. The high permease content of
the bacteria results in a sufficiently high
internal inducer concentration to overcome the
inhibitory effect of glucose.» in Cohn et
Horibata, 1959 |
||||||
|
«We investigated the kinetics of
f3-galactosidase formation by bacteria growing at low
inducer concentrations. Immediately upon the addition of
inducer, the rate of galactosidase synthesis per bacterium
rose linearly and continued in this way for a number of
generations. It was difficult to understand this result on
the assumption that each bacterium has about the same enzyme
content. We were able to show that this assumption does not
apply at low inducer concentrations. We discovered that at
the low inducer concentrations used in these experiments the
population consists essentially of individual bacteria that
are either making enzyme at full rate or not making it at
all. As the fraction of fully induced bacteria in the
population rises, there is an increase in the average rate
at which enzyme is produced. » Remarque: |
Fig3
in Novick et Weiner, 1957 --> Le premier résultat marquant est
que la synthèse des deux enzymes par chaque
bactérie est un phénomène
"tout ou rien". Soit la bactérie
synthètise les 2 enzymes de façon maximale,
soit pas du tout. Dans une population, la synthèse
globale n'est que la somme des synthèses
individuelles de chaque bactérie. fig
5 in Novick et Weiner, 1957
|
Novick A., Weiner M., 1957. Enzyme induction as an all-or-none phenomenon. Proc. Natl. Acad. Scie. USA 43, 553-566 (accès libre) |
* la durée d'une génération bactérienne est définie arbitrairement dans des conditions initiales par le temps de doublement de la population bactérienne de départ (mesurée par densité optique) divisé par ln2. Un chercheur qui explore la question de l'individualité: G.N. Amzallag (voir son article "Du sens de la variabilité" dans le même ouvrage "Génétiquement indéterminé"). Certains éléments de sa réflexion sont présentés sur la page sur les mutations. Remarques: - Cette manière de voir, qui met en valeur le rôle individuel de chaque molécule cytoplasmique (d'inducteur ou de perméase) est aussi une voie moderne de compréhension de la physiologie qui s'oppose à la vision statisticienne qui domine au XXème siècle (voir Au-delà de la cellule). |
|||||
|
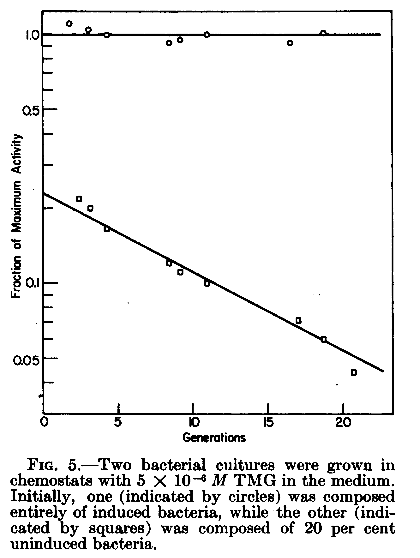
suite Discussion : «The model also explains the maintenance experiments. Thus it is quite possible that at the low concentrations of inducer used in maintenance experiments the threshold number of permease molecules needed to drive a bacterium to maximum induction may have a value greater than 1. Nevertheless, if the number of permease molecules at maximum is large compared to the threshold, there is a high probability that, on division of a fully induced bacterium, each daughter cell will receive a sufficient number of permease molecules to assure maximal induction by the maintenance concentration of inducer. Indeed, the fact that a maximally induced culture can be maintained maximally induced for many generations shows that the chance of a bacterium becoming uninduced under these conditions is very small. Were any uninduced organisms to appear, they would be selected for by their more rapid growth and would bring about a reduction in the rate of galactosidase synthesis of the culture. » |
||||||||
|
On s'éloigne de plus en plus de l'être vivant .... |
||||||||
|
Inhibition by glucose of the induced synthesis of the ß-galactoside-enzyme system of Eschericchia coli. Analysis of Maintenance, Melvin Cohn and Kengo Horibata, J Bacteriol. 1959 November; 78(5): 601&endash;612. article en libre accès pour le premier article
|
« SUMMARY An Escherichia coli clone growing in the presence of glucose, to which is added a ß-galactoside inducer, will not be induced to make either the galactoside-permease (Y) or ßgalactosidase (Z). The same clone, preinduced in the presence of inducer, will continue to make permease and enzyme, after glucose is added. These studies have shown that a given culture growing in a fixed medium can exist indefinitely in alternative steady states and that it can be shifted from one stable state to the other by transitory variations in the environment. A model which accounts for these findings is based upon the facts that (a) Y is induced by an internal galactoside whose accumulation is catalyzed by Y, and (b) glucose, in order to inhibit, enters by a distinct and independent route. In such a situation, the state of nonsynthesis is stable because the ß-galactoside cannot reach the site of Y synthesis and overcome the glucose inhibition in a cell which lacks Y. The state of synthesis is stable because the initial presence of Y provides the internal inducer necessary for the continued synthesis of Y in a glucose environment. The synthesis of Z simply mirrors the synthesis of Y since both entities are induced directly or indirectly by the same substance, namely the product of Y action.» |
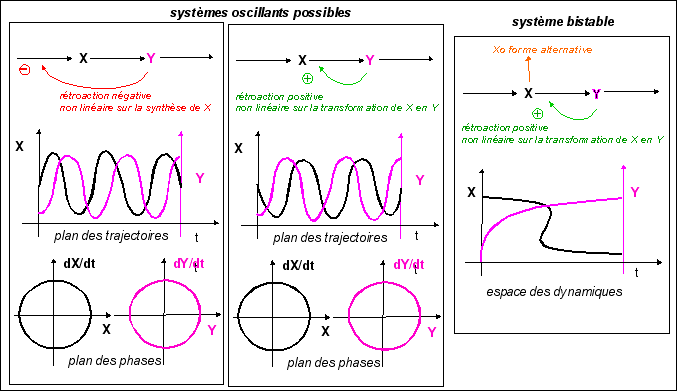
À partir de 1959 on va parler de bistabilité: présence de deux états stables chez un même clone de bactéries adaptées. Cette stabilité n'est pas contrôlée génétiquement de façon directe mais est due à la structure du réseau de régulation entourant la synthèse de ces enzymes. L'état stable se transmet de génération en génération, si les conditions de milieu restent stables. |
Remarque: |
|||||
|
|
||||||||
|
travaux de B.G. Hall des années 1975-1985 (travaux de cet auteur dans Pubmed dont un bon nombre sont en libre accès On the specificity of adaptive mutations, Hall BG., Genetics. 1997 Jan;145(1):39-44.PMID: 9017388 http://www.ncbi.nlm.nih.gov/pubmed/9017388 |
Depuis les années 1970 Barry G. Hall n'a cessé de faire un travail d'expérimentateur interprété dans un sens différent du mutationnisme ambiant. Il s'est intéressé depuis le début à l'opéron lactose. En 1975 il proposait déjà une redondance du réseau métabolique pour expliquer des changements de phénotypes (Lac-). Dans les années 1990 il a proposé à plusieurs reprises d'intégrer la notion de mutation adaptative qui s'oppose au dogme de la mutation apparue au hasard. Il publie toujours. Il s'oriente de plus en plus vers la phylogénie et la génomique. En travaux |
L 'homme végétal, Nissim Amzallag, Albin Michel, 2003, p 71-75 |
||||||
|
|
||||||||
|
Origin of Bistability in the lac Operon, M. Santillán, M. Mackey, E. Zeron, 2007, Biophysical Journal, Volume 92, Issue 11, Pages 3830-3842
On a pris l'habitude de désigner le gène par des minuscules et les polypeptides par des majuscules; ainsi lacZ (ou z) désigne le gène du polypeptide LacZ ou Z ou ßgalactosidase. |
Avec le développement des
études génétiques on découvrira
un troisième gène dans l'opéron lac.
Mais la connaissance encore très imparfaite du
rôle de la transacétylase (TRA) codée
fait que le modèle de bistabilité repose
encore de nos jours sur les deux seuls produits:
ßgalactosidase (Z) et perméase (Y). Les
modèles mathématiques qui montrent la
stabilité du système enzymatique ont
été passés en revue récemment
par Santillàn et Mackey (2008) par exemple, mais les
études continuent encore.
|
Quantitative approaches to the study of bistability in the lac operon of Escherichia coli, Moisés Santillán and Michael C. Mackey, J R Soc Interface. 2008 August 6; 5(supp1): S29&endash;S39, PMC2504340, http://www.ncbi.nlm.nih.gov/ pmc/articles/ PMC2504340/ |
||||||
|
|
||||||||
|
|
Deux limites qui me semblent
fondamentales pour de tous ces modèles: |
Ces études sont sans aucun doute
intéressantes et la direction originale prise par les
recherches en physiologie bactérienne est fortement
redevable aux disciplines d'origine de ses chercheurs:
physique et mathématique. C'est une richesse dont les
biologistes seraient fous de se passer mais ils ont le droit
de penser que leur objet d'étude, le vivant,
échappe à ce formalisme réducteur. |
||||||
|
|
||||||||
|
|
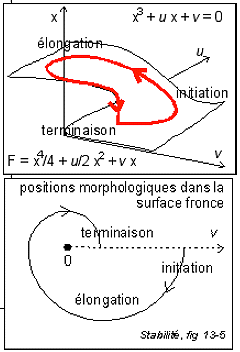
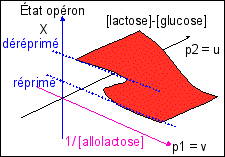
Le modèle continu le plus simple correspond à un cycle d'hystérésis avec conflit de deux attracteurs décrivant une surface de type "fronce" dans l'espace des dynamiques. |
|||||||
|
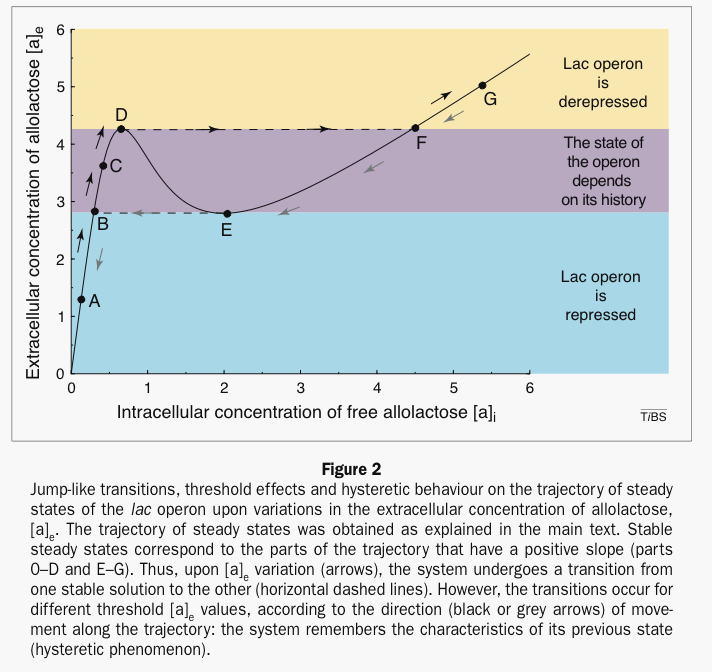
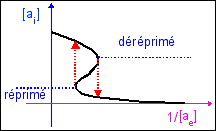
Michel Laurent et Nicolas Kellershohn présentent une courbe d'hystérésis, dans le plan des deux paramètres principaux (les teneurs en inducteur (allolactose) extra- [ae] et intracellulaires [ae]), dont voici l'allure.
|
Cette courbe est construite à partir des concentrations. Mais on peut imaginer facilement une courbe d'hysteresis similaire construite sur un modèle thomien de type fronce avec deux attracteurs. Deux états stables (expression/répression) pour la morphologie du système (les X ou sorties) avec hystérésis, et un paramètre de contrôle, la teneur en allolactose du milieu (inducteur extracellulaire). On peut ajouter un autre paramètre qui détermine le seuil d'apparition du phénomène d'hystérésis et qui pourrait être ici proportionnel à la différence entre la concentration en lactose et en glucose du milieu (puisque les bactéries sont sensibles à ces substrats).
|
Qu'apporte le modèle thomien par
rapport au modèle cinétique thermodynamique
(des équilibres chimiques) exprimé à
l'aide d'équations différentielles ? |
||||||
|
Pourtant, les postulats sur lesquels repose ce modèle sont fondamentalement différents de ceux des modèles chimiques. Dans le premier cas (chimie) l'état stable quantifie des interactions entre molécules sur lesquelles on fait un grand nombre d'approximations afin de pouvoir présenter des fonctions simples exprimant leur concentration et les relations entre elles. Dans le second cas (modèle thomien), les états stables (steady-state) correspondent à des attracteurs. La fonction modélise le comportement des bactéries et peu importent les molécules et leurs concentrations. C'est en quelque sorte un modèle "externe" qui s'oppose au modèle "interne" des chimistes. Mais dans un cas il y a mathématisation de la vie, dans l'autre mathématisation de la matière. Selon un autre point de vue on pourrait aussi dire que le modèle thomien est plutôt qualitatif alors que les modèles chimiques sont quantitatifs. Remarque: |
La démarche peut être poursuivie dans de nombreux cas où le modèle discontinu chimique s'avère être une impasse (ce qui est le cas de la majorité des situations métaboliques des eucaryotes). En effet, on suppose toujours un grand nombre de facteurs chimiques en relation, et le nombre d'équations différentielles devient vite ingérable. D'ailleurs, qui avait compris, et qui comprend encore aujourd'hui la démarche de Prigogine et Nicholis (voir ci-dessous) ? Alors que la démarche thomienne, qui part des morphologies des systèmes (et donc des données expérimentales), ne demande pas de calcul, mais donne une compréhension en profondeur des systèmes. Évidemment l'intérêt des modèles thomiens croîtra avec leur usage. Plus on s'appuiera sur ce formalisme, plus on démêlera l'écheveau des relations métaboliques que l'on a construit un peu aveuglément à partir de la chimie |
|||||||
|
|
||||||||
|
Une approche originale mathématique pour la thermodynamique .... |
||||||||
|
|
L'approche de Prigogine de l'école belge, à partir de la thermodynamique est difficile d'accès et aujourd'hui encore, même si ses théories sont souvent citées, il est impossible d'avoir accès gratuitement aux publications concernant ce modèle, qui date pourtant des années 70. L'ouvrage Self-organization in Nonequilibrium Systems. From dissipative Structures to Order through Fluctuations, Nicolis, Prigogine, 1977, reste non traduit (à ma connaissance) et protégé; l'opéron lactose y est décrit dans une quinzaine de pages, comme un système ouvert loin de l'équilibre mais présentant une organisation (structure) stable qui repose sur sa dynamique décrite 5 équations différentielles. Si je n'ai pas accès à ce modèle, je signale cependant que les articles de thermodynamique de l'Encyclopedia Universalis sont écrits par ces auteurs mais sans que les modèles soient vraiment présentés. |
Remarque: |
||||||
|
Remarque: |
« [...] Un milieu matériel peut accomplir un processus d'organisation désordre -> ordre, tout en restant en parfait accord avec le second principe de la thermodynamique [...]. On sait que ce même principe affirme aussi que l'état d'évolution le plus probable pour tout milieu isolé est l'état désordonné d'équilibre (maximum de l'entropie) et que cette propriété a souvent été invoquée en faveur d'une prétendue incompatibilité entre les lois d'évolution de la matière et celles de l'ordre biologique qui gouverne l'apparition de la vie. Les conclusions de Prigogine, et d'une façon générale l'ensemble des résultats obtenus par l'école thermodynamique de Bruxelles, ont permis d'éliminer cette incompatibilité apparente. L'interprétation nouvelle fait appel au mécanisme sous-jacent d'intervention des fluctuations. Au voisinage de l'équilibre, celles-ci disparaissent dès leur formation et peuvent donc être ignorées. Toutefois, dans la région non linéaire, certaines d'entre elles peuvent s'amplifier à proximité d'un premier état critique et, conformément à des lois stochastiques, venir perturber l'état macroscopique établi et le déstabiliser. Il en résulte un changement de branche ou bifurcation vers un nouvel état stable pouvant être plus structuré que le précédent devenu instable et, dès lors, éliminé. Des perturbations d'origine extérieure peuvent avoir le même effet. Les structures dynamiques ainsi formées sont essentiellement non isolables des contraintes extérieures imposées, ce qui détermine une distinction fondamentale avec les cristaux qui sont des structures statiques, donc d'équilibre. Comme les structures dynamiques exigent une dissipation constante d'énergie et de matière, Prigogine leur a donné le nom de structures dissipatives. Un premier exemple particulièrement édifiant par sa simplicité est celui de l'auto-organisation cellulaire de H. Bénard, lors de l'apparition de la convection libre dans une couche horizontale de fluide chauffé par le dessous. L'apparition des cellules de convection se produit à partir d'un seuil critique, c'est-à-dire d'un gradient thermique, et donc d'une activité productrice d'entropie suffisamment intense. (Article "Prigogine", Isabelle Stengers, EU 2010) |
|||||||
|
Les paragraphes suivants
sont extaits (ou inspirés) de
Multistabilité et
épigénèse dans les systèmes
biologiques, |
|
|||||||
|
En travaux |
||||||||
|
La plupart des modèles chimiques actuels intégrent cette dimension stochastique |
||||||||
|
|
||||||||
|
CONCLUSION pour un aperçu historique voir la cours de 1èreS
|
Les gènes impliqués dans la synthèse d'enzymes constituent une part importante du génome, mais il est maintenant déraisonnable d'imaginer qu'ils pourraient ainsi contrôler la totalité du métabolisme. Si le métabolisme lui-même a été considéré comme un réseau d'équilibres chimiques contrôlés par des enzymes, ce n'est plus le cas désormais. On s'intéresse maintenant au niveau structural (enzymes regroupées temporellement et spatialement en systèmes ou au sein de domaines cytoplasmiques... par exemple). Ce contrôle d'ordre structural peut être sous la dépendance de paramètres physiques. Le temps où l'on cherchait à tout expliquer en terme d'information est révolu. |
La raison malmenée, Nissim Amzallag, CNRS Éditions, 2002, p 35 « Contrairement à ce
qu'énoncent les paradigmes de la biologie
moléculaire, il n'y a pas de correspondance
étroite entre un gène et une fonction.
Même dans le cas le plus simple, celui des
réactions chimiques du métabolisme, le
contrôle opéré par les enzymes forme un
réseau, et non pas une chaîne de
réactions catalytiques. Or, les structures en
réseau connaissent plus d'un circuit fonctionnel, si
bien que la déficience d'un enzyme est
fréquemment contournée par d'autres voies
métaboliques. Il n'est donc possible de
déterminer une correspondance stricte entre
gène et fonction que pour des cas très
particuliers, ceux où pour une quelconque raison le
réseau de régulation disparaît. |
||||||
|
L'investigation du métabolisme, et principalement du métabolisme procaryote, tel qu'il a été mené jusqu'alors, à partir d'organismes sélectionnés comme "mutants", et qui sont en fait incapable d'adaptation, est une voie sans issue, même si elle continue de faire l'objet de la majorité des publications, y compris depuis que les physiciens et les mathématiciens s'intéressent aux réseaux métaboliques. |
Chez les eucaryotes, chaque gène du protéome, dans un environnement génétique et nucléaire donné, peut conduire à la synthèse d'un produit (ARN) qui, lui-même, conduit parfois à la synthèse d'un polypeptide. Les niveaux de transcription et de traduction peuvent être régulés de façon très globale par la synthèse de grands ARN en grand nombre ou au contraire faire l'objet d'une régulation strictement locale, liée aux environnements nucléaires qui dépendent de la vie de la cellule. Un même gène peut donner différents produits et chaque produit peut avoir des fonctions différentes au cours de la vie de l'organisme ou au niveau de tissus différents. |
|||||||
|
|
||||||||
|
3.2 -
L'expression aléatoire des
gènes
|
En travaux Source de ce chapitre: |
|||||||
|
|
L'opinion habituelle, qui cadre avec l'opposition kantienne entre déterminisme et liberté, considère que seul le déterminisme est scientifique, ce qui a pour conséquence de confondre "l'indéterminisme scientifique" avec un "déterminisme statistiquement indéterminé" : on ne peut prévoir ou retrouver les étapes d'un phénomène, non pas parce qu'il ne repose pas sur un déterminisme mais parce que les paramètres sont trop nombreux ou trop complexes pour qu'un résultat unique et prévisible en émerge. Le "hasard" kantien, qui est appelé ici "aléatoire", n'est donc pas un indéterminisme ontologique mais juste expérimental. On est en droit de considérer que
cette division kantienne est erronée et qu'en science
il existe un vrai indéterminisme qu'on ne peut pas
séparer de la liberté. |
|||||||
|
+ l'exemple de certains gènes lors du développement.... expression aléatoire (Paldi : Le problème de l'accessibilité de l'ADN eucaryote a été pris comme exemple par A. Paldi dans son article pour expliquer l'approche non déterministe du métabolisme que certains biologistes moléculaires proposent. |
||||||||
|
3.3
- Des gènes à tout
faire
|
En travaux |
|||||||
 .
.
|
3.4
- Voie thomienne: l'ADN, mémoire ou point stable des
dynamiques de synthèse
|
Source principale: |
|||||
|
Dans la
démarche de biologie théorique proposée
dans cette page, la compréhension de l'ADN est
inversée par rapport à la démarche
d'une théorie informationnelle; on ne pose pas comme
premier postulat que l'information va de l'ADN aux
protéines (ce qui vient ensuite), mais on s'efforce
d'abord de comprendre comment la cellule enregistre, dans
l'ADN, ses dynamiques; comment l'ADN est au cœur, comme
un centre organisateur, non pas qui dirige, mais qui est
comme un point stable des dynamiques de synthèse. Ne
peut-on envisager qu'il est vain de croire qu'il y a autre
chose dans l'ADN qu'une trace mémorisée (et
transmissible ? ... au moins partiellement) des
synthèses de la cellule (rien sur la forme, rien sur
le mouvement...).
|
||||||
|
3.41 L'ADN est répliqué contrairement aux protéines |
||||||
|
|
Les caractéristiques chimiques simplifiées de l'ADN (voir cours de seconde) sont supposées connues. |
On dit souvent que les acides
nucléiques ne peuvent former seuls de grands
assemblages macromoléculaires alors que les
protéines le peuvent. C'est relativement faux
puisque, lorsque l'ADN n'est que peu entouré de
protéines (bactéries,
certains protozoaires, certains noyaux de
spermatozoïdes...), on observe
des phases dites
cholestériques de filaments plus ou
moins orientés. Ces phases sont de même nature
que celles observées dans le cas des fibres de
cellulose ou de collagène... (voir les travaux
d'Yves Bouligand (EPHE, voir articles dans
E.U.: phases mésomorphes,
cristaux liquides) ou de
Françoise LIVOLANT (Laboratoire
de Physique des Solides, Université Paris Sud,
Orsay); voir la page
sur l'ADN condensé.
|
||||
|
l'ADN
lors des cycles cellulaires |
La réplication (ou duplication) de l'ADN est catalysée par de nombreuses enzymes mais le principal complexe enzymatique est l'ADNpolymérase (voir complément cours de 1èreS). La réplication de l'ADN est dite semi-conservative car, dans chaque molécule issue de la réplication, un brin est néosynthétisé, et un brin est ancien.
|
Chez les procaryotes il semblerait que la synthèse d'ADN puisse avoir lieu en permanence, même pendant une division. Chez les procaryotes il pourrait y avoir habituellement plusieurs exemplaires de la molécule d'ADN unique dans chaque cellule (6 pour Escherichia coli). |
Chez les eucaryotes la réplication se fait pendant la phase S (dite de SYNTHÈSE) du cycle cellulaire, c'est-à-dire pendant l'interphase, phase de repos nucléaire. On considère que chaque molécule d'ADN est totalement répliquée et une seule fois avant chaque division chez les eucaryotes. Il y aurait une molécule d'ADN par chromatide de chaque chromosome. |
|||
|
|
||||||
|
|
Il est donc probable que la synthèse de l'ADN ne soit pas seulement un mécanisme enzymatique (discontinu avec progression) mais qu'il dépende d'une dynamique plus catastrophique (au sens de Thom et donc un phénomène CONTINU) qui engendrerait ce passage d'une à deux molécules par séparation des brins et synthèse de brins complémentaires. On sait arrêter la synthèse de l'ADN mais dans ce cas la cellule ne se divise pas et sa dynamique est brisée, ce qui est aussi un argument en faveur d'une explication continue. |
Si l'on reprend les dynamiques élémentaires trouvées par René Thom (tableau de la page du cours de 1èreS) on a l'embarras du choix pour générer une duplication à partir du moment où l'on considère un conflit entre deux attracteurs (et donc à partir de la fronce). Mais René Thom n'a pas proposé de modèle spécifique à la réplication de l'ADN. Il intègre cette duplication dans sa blastula physiologique (voir annexe cours 1èreS) avec les duplications de cellules (mitose) et duplications d'organites (le texte le plus abouti est celui qui se trouve dans Esquisse d'une sémiophysique (Paris 1988). Le livre demande à être lu dans son ensemble même si cette question est particulièrement traitée dans la partie F (Le problème de la duplication des cycles plans, pp 85s). Je ne maîtrise pas encore sa formulation au point d'en faire une explication à destination des élèves... mais j'y travaille. |
Enfin, il n'est pas impossible que la réplication de l'ADN bactérien et de l'ADN eucaryote ne soient pas tout à fait équivalent du point de vue des dynamiques étant donné la formation du chromosome des eucaryotes qui est une structure tout à fait originale.
Une fois encore, pour mieux
comprendre la différence entre modèles
continus et discrets en biologie, |
|||
|
Si l'on reprend les idées exprimées dans le cours de 1èreS sur la différence de temps constitutif des dynamiques de synthèse des protéines et des dynamiques qui conduisent à la synthèse de l'ADN on aborde maintenant une autre façon de voir l'ADN, non plus comme substrat mais comme produit. L'ADN mémoire est une molécule synthétisée sur le long terme par des processus dont on n'a pas forcément pris la bonne mesure. Dans ce cas il n'est pas étonnant que l'on ne puisse suivre que la duplication de l'ADN et sa pseudo-stabilité si l'on n'a pas la bonne échelle de temps puisqu'elle ne serait pas identique pour tous les organismes. Je précise qu'il ne s'agit pas de chercher une vitesse de synthèse (qui peut être réalisée in vitro avec un temps lié à la cinétique chimique enzymatique à partir des composants extraits) mais bien de rechercher les conditions d'apparition de la bifurcation correspondant à la synthèse d'ADN dans le temps héréditaire ou plutôt divisionnel (lié au phénomène de division). |
||||||
|
3.42 L'ADN est transcrit de façon discontinue |
||||||
|
Dans la théorie de l'information
génétique on a postulé que la
transcription était l'étape clé du
contrôle des gènes moléculaires
(voir
cours 1ère S). De
très nombreux modèles actuels de
réseaux génomiques sont basés sur cette
hypothèse. |
Transcriptional Maps of 10 Human Chromosomes at 5-Nucleotide Resolution, Cheng J. et al. Science 308, 1149-1154 (2005). accessible librement sur internet à l'adresse: http://www.euchromatin.com/ ChengJ01.htm) |
|||||
|
Qu'est-ce qu'un
gène ? |
« Au lieu d'avoir des gènes discrets transcrits scrupuleusement en ARN identiques, la transcription convertirait de nombreux segments du génome en une masse envahissante de rubans d'ARN de différentes longueurs. Les rubans peuvent être transcrits à partir des deux brins de l'ADN contrairement à ce qui est habituellement cru. Certains de ces transcrits viennent de régions déjà identifiées comme contenant des gènes codant pour des protéines. Mais beaucoup de viennent pas de ces régions. "C'est quelque peu révolutionnaire", dit Phillip Kapranov, le collègue de Gingeras. "Nous avons compris que le génome est plein de transcrits chevauchants" ». |
En travaux |
||||
|
- enfin, le troisième modèle, déterministe et/ou indéterministe mais continu, où l'ADN est un point stable des dynamiques de synthèse. Ce modèle reste à construire, il n'a été que suggéré par René Thom. |
||||||
|
|
(in Esquisse d'une sémiophysique, fig 4.29) |
l'ADN est au centre d'une structure cinétique cyclique où les protéines sont synthétisées au bord de la spirale en rotation (attention il ne s'agit pas d'un modèle dans l'espace euclidien mais dans l'espace des phases...voir page sur les modèles thomiens). Cependant ce modèle repose grandement sur la compréhension du cytoplasme comme une phase fluide ce qui est franchement erroné (voir page sur la cellule).
Il faut donc chercher un modèle plus moderne et plus élaboré |
||||
|
Pour ce qui est de la transcription, un modèle thomien (fronce) est présenté sur la page sur les enzymes |
|
|||||
|
3.43 L'ADN est-il stable ? Contrairement aux protéines l'ADN n'est pas synthétisé en permanence, utilisé, puis dégradé, il est plutôt multiplié et partagé. mais cela ne veut pas dire qu'il n'est pas modifié en permanence, même si la quantité d'ADN reste plus ou moins fixe dans la cellule.... |
||||||
|
* l'ADN est
fragile |
Les chromosomes aussi sont fragiles, et même s'il ne contiennent qu'environ 50% d'ADN, la présence de très nombreux sites fragiles que l'on commence à recenser, est un argument en faveur d'un remaniement permanent de ces structures. voir chromosomes |
|||||
|
Pour un survol de la partie historique, voir la partie 1.1 de la page du cours de 1èreS. |
* dans
la théorie de l'information génétique
on pense que c'est la séquence de l'ADN qui a un
sens... dans une théorie morphodynamique, taille,
forme, organisation spatiale en sous-domaines, origine,
emplacement,..., sont autant d'éléments qui
donnent des sens. |
Quelques postulats de la biologie
moléculaire remis en cause: voir page sur l'identité biologique de l'homme |
||||
|
ADN grand voyageur, brève d'Olivier Donnars, La Recherche, 413, nov 2007, 14-15 |
|
|||||
|
Equipe travaillant sur les réversions sexuelles de crustacés isopodes suite à l'infection par Wolbachia: Laboratoire de Génétique et Biologie des Populations de Crustacés (UMR 6556) http://ecoevol.labo.univ-poitiers. fr/spip.php? rubrique31 voir l'article « la bactérie qui rend femelle» La Recherche, n° 293 décembre 1996 http://ecoevol.labo.univ-poitiers.fr/ IMG/pdf/articles_ cloportes_ 4-2.pdf |
Certains organismes eucaryotes pluricellulaires (plantes et animaux), qualifiés de symbiotes obligatoires, nécessitent une infection bactérienne pour que leur lignée ovocytaire ou leurs graines se développent. D'autres symbioses, comme celle de bactéries du genre Wolbachia avec de nombreux arthropodes et nématodes, facultatives, interviendraient dans le contrôle de la fécondité des hôtes (l'infection conduisant parfois à une réversion sexuelle des femelles en mâles, voir références dans la colonne ci-contre <----). On se doutait que l'interaction hôte-symbiote était très forte mais on a découvert, dans le cas de la bactérie symbiotique Wolbachia pipientis, que la bactérie laissait des traces de son passage dans le génome même des cellules eucaryotes, mêmes celles qui ne contenaient plus d'endosymbiotes (voir résumé de l'article ci-contre --->). |
|
Widespread Lateral Gene Transfer from Intracellular Bacteria to Multicellular Eukaryotes, Julie C. Dunning Hotopp et al., Science, 317, 21 sept 2007, 1753-1756 |
|||
|
Mais les résultats
génétiques obtenus sont prudemment à
mettre en regard avec les études physiologiques et
écologiques menées par ailleurs pour
éviter encore une fois de retomber dans le tout
génétique (comme le
suggère une fois encore la conclusion conventionnelle
du résumé ci-contre) :
pour moi l'ADN étranger est la marque des fonctions
nouvelles et non la cause.
|
||||||