Plan de cette page
Annexes
* Des caractères des individus et des espèces
* Remarques sur le programme (et justification du plan suivi)
L'exemple qui nous est demandé d'étudier cette année est le développement des plantes, ce qui nous permettra des comparaisons avec le développement animal, traité en classe de seconde (voir cours de seconde). Le développement animal était aussi survolé dans cette même partie de l'ancien programme de 1èreS; si vous trouvez d'anciens manuels scolaires, cela peut aussi être une source de documents.
Sources :
Le cycle cellulaire chez les animaux et les végétaux,
Jean Clos, Marc Coumans et Yves Muller, Biologie-Géologie
(revue de l'APBG), n°3-2002, p497-564
Biologie végétale, tome 2 : Organisation
végétative, D. Robert et A.M. Catesson, Doin, 1990
Biologie moléculaire de la cellule, Alberts et al., 1994,
Médecine-Sciences Flammarion
Biologie du Développement, S.F. Gilbert, 1996, De Boeck
Université
Microbiologie, Prescott, Harley et Klein, 1995, De Boeck
Université
Les unicellulaires, et les procaryotes, se divisent pour
se reproduire, après une phase de croissance
très générale. Ils bouclent ainsi ce qu'on
appelle leur cycle cellulaire.
Les pluricellulaires, donc eucaryotes divisent leurs cellules pour
se développer (et croître), se renouveller
(renouvellement cellulaire) et se reproduire. Le cycle
cellulaire de leurs cellules n'est donc qu'un des
éléments du cycle de développement (ou
cycle de vie pour l'organisme entier) qui contient aussi des
phases de croissance, mais encore des phases de
différenciation et de sénescence, que nous verrons dans
le chapitre suivant.
Le cycle cellulaire comprend les
événements situés entre deux divisions.
Il comprend une phase de croissance cellulaire de durée
variable, pendant laquelle l'ADN est répliqué, et une
phase de division de durée fixe (20 min) chez la
bactérie Escherichia coli, qui répartit l'ADN dans les
deux cellules filles qui se séparent.
Il commence donc, pour une cellule, lors de sa séparation d'avec sa cellule sœur; et finit lors de sa séparation en deux cellules filles. Le cycle cellulaire commence par un allongement de la cellule, sans augmentation de diamètre, qui atteint ainsi 2 fois sa longueur initiale : c'est la croissance cellulaire. Comme il n'y a pas d'augmentation de diamètre on peut penser que la masse initiale double pendant cette croissance étant donné que la bactérie peut être assimilée à un cylindre de diamètre constant. Pendant cette croissance l'ADN est dupliqué et de nombreuses protéines dites de division sont synthétisées. Si l'on bloque la réplication de l'ADN on empêche la division et la cellule s'allonge démesurément en formant un long filament. La division cellulaire commence presque toujours, quelque soient les conditions de culture de la bactérie et donc quelque soit la durée du cycle cellulaire, 20 minutes avant la fin de la réplication.
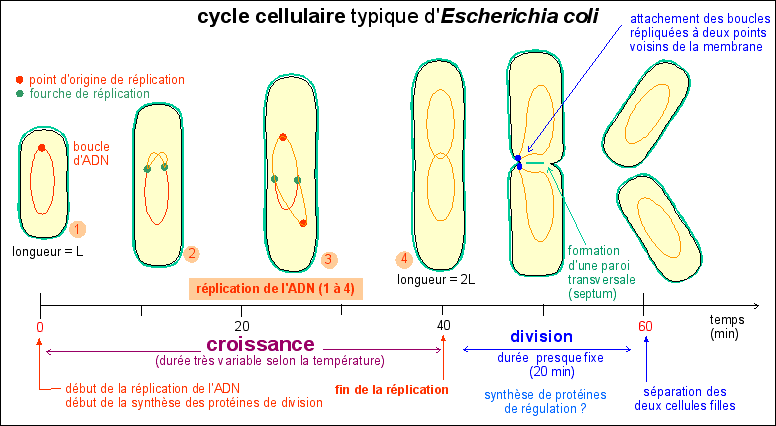
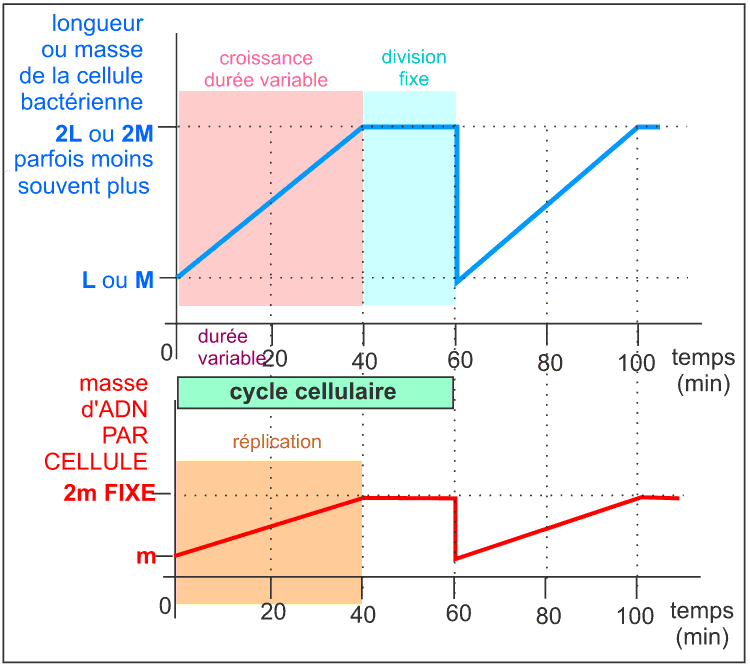
![]() Le cycle cellulaire - d'une durée de 60 min - d'une souche
d'Escherichia coli à croissance lente
Le cycle cellulaire - d'une durée de 60 min - d'une souche
d'Escherichia coli à croissance lente
Remarque:
Le cycle cellulaire ne se superpose pas au cycle de vie
bactérien car ne nombreuses bactéries peuvent sporuler,
c'est-à-dire former une spore résistante, qui
leur permet notamment de résister dans des milieux
extrêmes et d'être transportées par l'air sur de
très longues distances.
De plus il existe des conjugaisons bactériennes qui
sont des échanges d'ADN entre bactéries dites
recombinantes.
La réplication de l'ADN est de loin le mécanisme le
mieux connu de tout le cycle cellulaire des Procaryotes. Elle a
particulièrement été étudiée chez
Escherichia coli. La division est fort mal connue.
L'expérience historique de Meselsohn et Stahl a permis de
préciser le type de mécanisme de réplication de
l'ADN.
|
|
|||||||||||||||||||||||||
|
Escherichia coli est cultivée pendant de
nombreuses générations (= durée d'un
cycle cellulaire puisqu'une génération
sépare une cellule mère de sa cellule fille)
sur un milieu de culture contenant des
désoxyribonucléotides marqués à
l'15N (témoin 1). Cette souche de
départ est transférée pendant un
génération sur un milieu contenant uniquement
des désoxyribonucléotides marqués
à l'14N (expérience 1). Enfin on
transfère des bactéries issues de cette
culture à nouveau sur un milieu contenant uniquement
des désoxyribonucléotides marqués
à l'15N (expérience 2). |
|||||||||||||||||||||||||
Interprétation: |
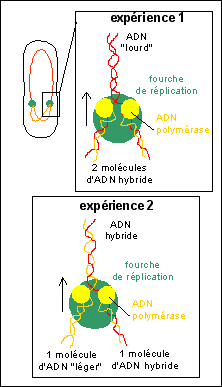 Une illustration de l'interprétation du mécanisme de réplication semi-conservative de l'ADN dans l'expérience de Meselson et Stahl. |
||||||||||||||||||||||||
La réplication de l'ADN est une synthèse de deux molécules d'ADN identiques à partir d'une molécule d'ADN. Chaque molécule d'ADN fille est composée d'un brin ancien et d'un brin nouveau complémentaire du brin ancien; on dit que la réplication est semi-conservative. La réplication de l'ADN est catalysée par un gros complexe enzymatique contenant plus de 20 enzymes mais surtout l'ADN polymérase. La réplication commence en un point origine et se poursuit en général dans deux sens opposés au niveau de fourches de réplication. Chez les eucaryotes, du fait de la longueur du génome, elle commence en de multiples points origine simultanément. (Il faudrait plus de 500 h à raison de 50 nucléotides à la seconde pour répliquer un seul chromosome humain à partir d'un seul point origine).
L'ADN polymérase nécessite une molécule d'ADN à répliquer, des désoxyribonucléotides, de l'énergie (ATP) et un petit fragment d'ARN complémentaire d'une petite séquence de l'ADN à répliquer (amorce). Il existe plusieurs ADN polymérases chez les bactéries (au moins 3) et plus encore chez les eucaryotes. La réplication se déroule avec une fidélité excellente: chez E. coli on estime le nombre d'erreurs de copie à 1 pour 109 à 1010 nucléotides, ce qui fait, pour un génome évalué à 4,7.106 paires de bases, une erreur pour 1.000 à 10.000 réplications. Il existe des systèmes de réparation de l'ADN qui utilisent aussi une ADN polymérase. La vitesse de polymérisation (ajout des désoxyribonucléotides pour former le nouveau brin) est de l'ordre de 20 à 1.000 nucléotides par seconde chez E. coli.
Le problème du déterminisme du cycle cellulaire est aussi celui du lien entre la croissance cellulaire et la réplication de l'ADN.... il est loin d'être résolu. En tout il ne sera pas traité dans ces pages.
Le cycle cellulaire des eucaryotes à surtout été étudié chez la levure de bière. Les données obtenues chez les cellules en culture et les cellules embryonnaires sont les plus nombreuses pour les animaux. Selon l'organisme, l'âge de la cellule, et le type cellulaire, on a des différences fondamentales. Seuls quelques points communs seront présentés ici. En tout cas il est clair que pour un pluricellulaire la notion de cycle de développement, qui se réfère à l'individu, est plus adéquat que la notion de cycle cellulaire, qui ne rend compte que de la vie d'une seule cellule.
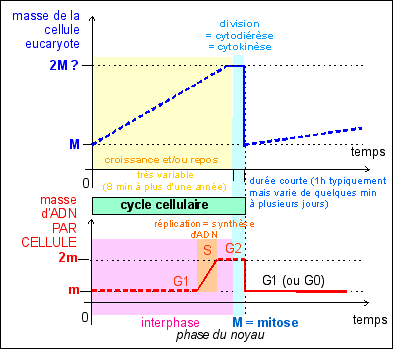 |
Le cycle cellulaire des cellules eucaryotes est beaucoup plus variable que celui des cellules procaryotes. Deux phases sont toujours observées:
|
En interphase le noyau est au repos du point de vue de la division mais pas de la synthèse car c'est en phase S de l'interphase qu'a lieu la réplication de l'ADN qui précède toute division (sauf dans le cas de la deuxième division de la méiose, voir cours de terminale).
L'expérience de Taylor met en évidence une caractéristique essentielle de cette phase de réplication: c'est une réplication semi-conservative. Elle fait appel aux mêmes mécanismes que la réplication chez les procaryotes. Les molécules d'ADN polymérases et les nombreuses enzymes associées sont bien moins connues que chez les procaryotes.
|
|
|
|
Les cellules eucaryotes cultivées in vitro sont des fragments racinaires de Bellevalia romana, une plante de la famille du Lys. La réplication de l'ADN est suivie à l'aide de thymidine tritiée (la thymidine est le nucléoside (thymine + sucre)) marquée par le tritium (3H ou 3T) isotope lourd et radioactif de l'hydrogène. L'incorporation du désoxyribonucléotide radioactif est suivi par des autoradiographies qui sont des expositions de préparations cellulaires à une plaque ou un film photographique. Les composés argentés de l'émulsion photographique précipitent lorsqu'ils rencontrent un électron émis par radioactivité ß- par les atomes de 3H incorporés dans le thymidine radioactive. Les grains d'argent forment des points noirs que le développement révèle. L'emplacement des points noirs indique donc de façon approximative la position des molécules de thymidine tritiée de la préparation cellulaire. Les temps d'exposition des émulsions photographiques sont bien sûr beaucoup plus longs que pour une impression par les photons. |
|
|
Les racines de Bellevalia romana, sont cultivées pendant la durée d'un cycle cellulaire sur un milieu "chaud", c'est-à-dire contenant de la thymidine tritiée. Les cellules en métaphase de mitose sont repérées et certaines sont autoradiographiées. L'aspect d'un chromosome est représenté ci-dessous à gauche. Certaines de ces cellules sont replacées immédiatement sur un milieu "froid", c'est-à-dire dépourvu de thymidine radioactive , pendant la durée d'un deuxième cycle cellulaire. L'aspect d'un chromosome d'une autoradiographie d'une cellule en prophase de mitose est présentée ci-dessous à droite. |
|
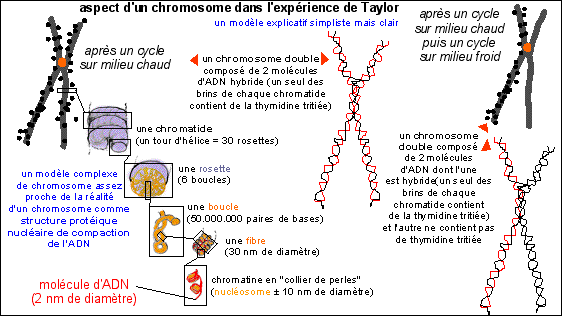 Illustration de l'interprétation de l'expérience de Taylor (en orange: le centromère (qui n'est pas visible); les points noirs correspondent aux grains d'argent de l'autoradiographie) (schémas d'après Principe de Biochimie, Lehninger et al, 1994, Flammarion-Médecine-Sciences) |
Explication de
l'expérience: Les chromatides radioactives sont composées d'une molécule d'ADN hybride. C'est-à-dire que la molécule d'ADN hybride à incorporé dans UN de ses brins, le brin nouvellement formé, de la thymidine tritiée. Cette incorporation n'a lieu que pendant la phase S du cycle cellulaire (synthèse de l'ADN) et ne touche donc que les cellules qui étaient dans cette phase de l'interphase peu après le début de la mise en culture. Lorsque les cellules sont retirées du milieu chaud et observées, seules celles qui se trouvent maintenant en métaphase peuvent présenter de tels chromosomes. Le fait que les deux chromatides, issues de la réplication de l'ADN en phase S de l'interphase, sont TOUTES LES DEUX hybrides (c'est-à-dire composées d'un brin ancien, non radioactif et d'un brin nouveau ayant incorporé de la thymidine tritiée radioactive) peut être expliqué par un mécanisme semi-conservatif de la réplication de l'ADN, comme chez les Procaryotes. |
Remarque:
les molécules d'ADN dans le noyau en interphase ne sont pas
libres et flottant dans le nucléoplasme mais sont à
l'état de chromatine. La
chromatine a un aspect en "collier de perles" (voir
l'illustration ci-dessus), est colorable en
ME (par le
tétroxyde d'Osmium) et comporte
un filament d'ADN et des protéines (des
histones) regroupées en petits cylindres (les
nucléosomes). Cette structure ne semble
pas empêcher la réplication et les nucléosomes
semblent être aussi présents dans le chromosome qui
résulterait alors de la compaction de la
chromatine (voir de nouveau
l'illustration ci-dessus qui n'est cependant qu'un modèle -
voir cours de
seconde pour quelques remarques à
ce sujet).
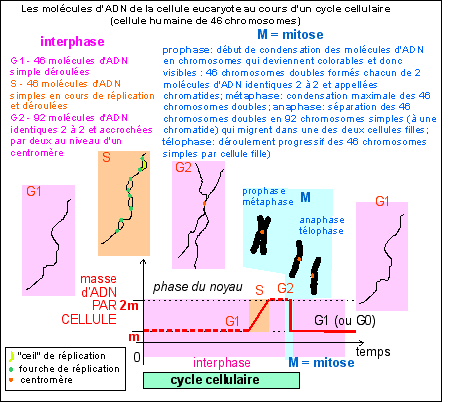 |
Place de la réplication dans le cycle cellule d'une cellule eucaryote et aspect très schématique des chromosomes au cours des différentes phases nucléaires.
Le "piège" pour un élève inattentif vient de ce que certains auteurs qualifient, improprement, de chromosome, tout filament d'ADN de la cellule. Ce mot doit être réservé aux structures colorables et donc composées d'ADN et de PROTÉINES de la cellule en phase M du noyau (voir ancien cours de spécialité de terminale). |
Les étapes de la mitose sont
classiquement regroupées en 4 phases qui présentent
bien sûr des limites parfois floues ou chevauchantes:
* la prophase où la chromatine se condense et forme les
chromosomes qui deviennent COLORABLES et VISIBLES en
microscopie optique; ce sont des chromosomes doubles (à deux
chromatides issues de la réplication en phase S de
l'interphase); l'enveloppe nucléaire se
désagrège par fragmentation des citernes du
réticulum qui entourent le nucléoplasme ;
* la métaphase où les chromosomes MIGRENT car
ils sont TIRÉS par les microtubules kinétochoriens et
se disposent de part et d'autre du plan équatorial de la
cellule en division; ce plan décrit de façon
précise l'orientation des deux cellules filles
(il est
matérialisé par un anneau ou un disque de filaments
d'actine, voir partie b suivante); si la
cellule est étirée, ce plan peut diviser la cellule
longitudinalement ou transversalement ce qui donnera une division en
épaisseur ou en longueur, déterminante pour la
croissance des cellules végétales à paroi par
exemple; entre les deux pôles de division, situés
symétriquement par rapport au plan équatorial, se
mettent en place des microtubules qualifiés de fusoriaux car
ils dessinent un fuseau de division;
* l'anaphase où les chromosomes doubles SE SCINDENT au
niveau des centromères, chaque chromosome fils (composé
d'une chromatide) est TIRÉ vers l'un des pôles de la
cellule par ses microtubules kinétochoriens qui glissent le
long des microtubules du fuseau de division;
* la télophase où les chromosomes se
déroulent et les filaments d'ADN redeviennent NON COLORABLES
sous forme de chromatine; l'enveloppe nucléaire se met
à nouveau en place autour du nucléoplasme.
Remarque:
toutes les "figures de mitose" ou "cellules en mitose"
magnifiques de vos livres scolaires photographiées au
microscope optique sont des COLORATIONS SPÉCIFIQUES des
seuls CHROMOSOMES et, si la cellule vous semble VIDE, c'est qu'elle
est TRANSLUCIDE et NON COLORÉE artificiellement comme le sont
les chromosomes (rappelez-vous l'étymologie de chromosome:
chromo = couleur et soma = corps: des corps
colorables). Vous pouvez comparer ces photos avec les coupes
ultrafines fixées et colorées (au tétroxyde
d'osmium, colorant opaque aux électrons) observées au
microscope électronique: vous verrez alors le
cytoplasme nettement plus encombré; tout en sachant que,
là encore, on ne colore, avec le tétroxyde d'osmium,
que certaines structures et non pas toutes... (par exemple: Bordas p
98, 103; Nathan p 105).
Plus difficiles à observer que les chromosomes, les
composants cytoplasmiques sont de mieux en mieux connus, du fait des
progrès des techniques d'observation et de marquage
spécifique. Les films pris au microscope à contraste
interférentiel, ont laissé désormais la place
aux films présentant un marquage immunologique par
plusieurs substances fluorescentes de couleurs différentes
qui permettent in suivi IN VIVO des protéines comme la
tubuline (en VERT le plus souvent), composant
des microtubules, de l'actine (en ROUGE le plus
souvent), composant de certains filaments fins de la cellule,
ou encore des histones (en BLEU le plus
souvent), composant de la chromatine (Nathan p
115; voir par exemple le CDRom qui accompagne la nouvelle
édition (2004) de Biologie Moléculaire de la Cellule,
Alberts et al., Médecine-Sciences-Flammarion... de très
nombreux films au format ".mov" sont disponibles... et copiables pour
être visionnées avec Quick-Time en
vidéoprojection.... une merveille de souplesse
pédagogique). Cette vision est cependant plus chimique
que structurale et il faut se garder d'abandonner les anciennes
méthodes. Les nouvelles méthodes doivent
compléter les anciennes.
Une coloration spécifique (rouge) existe pour les microtubules
en microscopie optique (voir par exemple Bordas p 102-103).
Les microtubules (cylindres creux et rectilignes de 25 nm de diamètre) et les filaments d'actine (microfilaments flexibles d'environ 5 à 9 nm de diamètre) constituent un cytosquelette dynamique. Il existe aussi des filaments intermédiaires (d'environ 10 nm de diamètre), par exemple ceux qui constituent un réseau dense situé contre la membrane interne de l'enveloppe nucléaire et que l'on qualifie de lamina nucléaire.
Les mitochondries, les chloroplastes, l'appareil de Golgi, le réticulum endoplasmique sont des structures incapables de se régénérer à partir de leurs composants, ils doivent être transmis par hérédité cytoplasmique directe. Les plus gros se fragmentent (RE, Golgi, chondriome (voir remarque ci-dessous)), le plus petits se répartissent après une phase de synthèse-multiplication (mitochondries ? chloroplastes ?). Une phase de synthèse membranaire intense fait suite à la cytodiérèse.
Remarques:
* Des données cytologiques récentes tendent à
montrer que le chondriome (ensemble des mitochondries d'une cellule)
pourrait notamment être formé d'un réseau de
tubules dynamiques anastomosées comme l'est le REL. C'est par
exemple le cas chez la levure.
* Les microtubules forment un réseau qui rayonnent à
partir d'une structure particulière qualifiée de
"centre organisateur des microtubules" qui, chez les cellules
animales, comporte deux petits cylindres protéiques creux
disposés perpendiculairement l'un à l'autre que l'on
appelle centrioles. Le centre organisateur des cellules
animales comportant deux centrioles est appelé
centrosome.
* Les microtubules sont des structures dynamiques qui s'allongent ou
raccourcissent très rapidement et continuellement. Les
microtubules accrochés au centromère des chromosomes
(fibres kinétochoriennes accrochés par le
kinétochore à des parties spécifiques du
centromère) se raccourcissent ainsi en tirant les chromosomes
fils vers les pôles où sont situés le centre
organisateur ou le centriole dans la cellule animale.
* De protéines motrices semblent pouvoir se déplacer le
long des microtubules en transportant de grosses molécules,
des particules protéiques ou des organites.
* Les cils et flagelles sont des composés complexes de
microtubules associés à des protéiques
particulières.
* Le début de la division cellulaire peut se repérer
dans sa phase la plus précoce par le changement d'organisation
du réseau des filaments d'actine du cytosquelette, ce qui se
fait en fin de phase G2 pour la cellule des plantes, avant que les
chromosomes se condensent et marquent le début de la
prophase.
 |
La division d'une cellule animale (phase M du cycle cellulaire) comprenant typiquement :
Les événements chromosomiques, voir ci-dessus, n'ont pas été détaillés. Le cytoplasme, moins connu, très encombré
et structuré n'a pas non plus été
détaillé: |
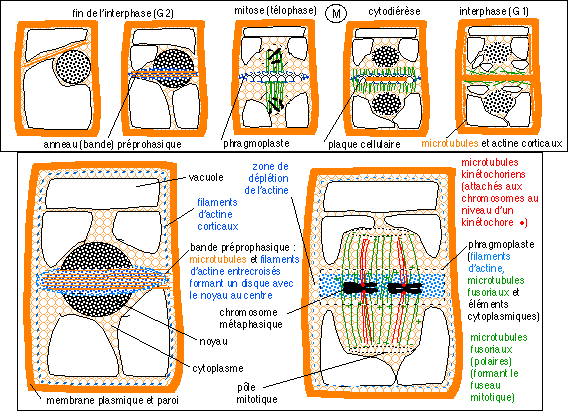 |
Division cellulaire d'une cellule de plante (phase M comprenant la mitose et la cytodiérèse). La fin de l'interphase précédant cette
division et le début de l'interphase qui la suit ont
été représentées. On retiendra le rôle essentiel des microtubules et des filaments d'actine dans les mouvements déterminant le plan de division, la mitose et la cytodiérèse. |
Remarques:
|
Procaryotes |
|
|
|
|
une cellule mère se divise et donne naissance à deux cellules filles |
La cellule procaryote se divise, se transforme en spore ou meurt. La division des Procaryotes est une division binaire ou division par scissiparité. La boucle d'ADN se duplique en deux boucles qui se sépareraient par allongement de la cellule du fait que chaque boucle (l'ancienne et la nouvelle) serait accrochées à la membrane en un point différent. Les ribosomes et autres éléments cytoplasmiques seraient plus ou moins également répartis dans les cellules filles. |
||
|
Protistes |
La cellule eucaryote présente des périodes de division courtes (environ 1h) et des périodes de repos, plus longues. La division cellulaire comprend deux étapes qui se chevauchent plus ou moins : la division nucléaire ou mitose et la division cytoplasmique ou cytodiérèse. |
Les modalités sont extrêmement variées: on parle de mitose atypique pour désigner les divisions nucléaires originales (certaines cellules séparent leur chromosomes en étirant l'enveloppe nucléaire, qui reste intacte, les centromères étant accrochées à sa membrane interne; des fuseaux de microtubules traversent souvent le noyau sans que l'enveloppe nucléaire ne se désagrège; chez les levures et les Diatomées, l'enveloppe nucléaire reste aussi intacte, le fuseau de division se forme à l'intérieur du noyau qui s'allonge et les chromosomes sont tirés par un seul microtubule kinétochorien...). |
|
|
Mycètes |
Lors de la mitose, l'enveloppe nucléaire se désagrège et un fuseau mitotique se forme; les chromosomes sont tirés par les microtubules kinétochoriens. Le centre organisateur des microtubules n'est bien individualisé (sous forme d'un centrosome contenant deux centrioles) que chez les animaux. |
||
|
Animaux |
|||
|
Végétaux |
|||
|
matériel génétique nucléaire |
- dupliqué avant la division
(sauf exception) |
||
|
matériel génétique mitochondrial ou chloroplastique |
transmis avec l'organite qui habituellement se dupliquent et se répartissent de façon à peu près égale entre les deux cellules filles |
||
|
cytoplasme et organites cytoplasmique |
le cytoplasme, avec ses organites et après duplication d'un bon nombre de ceux-ci, est réparti de façon équitable entre les deux cellules filles |
||
|
origine de la cellule mère |
une cellule mère peut être une jeune cellule en fin de division (par exemple lors de la multiplication cellulaire du développement embryonnaire ou une cellule bactérienne...) mais la cellule mère peut aussi est une cellule quiescente qui est stimulée brusquement u encore une cellule différenciée qui peut se diviser en se dédifférenciant ou non. |
||
|
devenir des cellules filles |
Certaines cellules filles meurent, d'autres se divisent presque à la suite, d'autres enfin entrent en quiescence... |
||
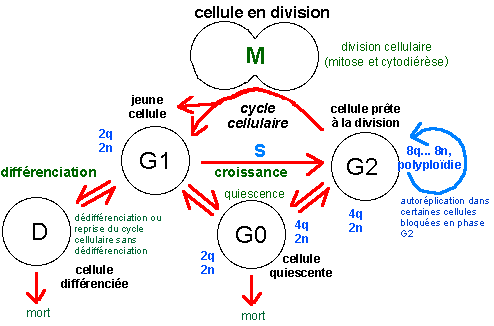
Aucune voie n'est complètement fermée tant que la cellule est vivante. Il n'en reste pas moins que certaines voies sont nettement plus empruntées que d'autres. Dans la phase de développement embryonnaire la plupart des cellules nouvellement formées se divisent à nouveau, même si parfois la division est inégale dans le sens où seule l'une des deux cellules filles garde son pouvoir mitogène, l'autre se différenciant (on parle de division formative par opposition à une division proliférative, où les deux cellules filles gardent leur pouvoir mitogène). Les division chez un organismes adulte sont nettement plus localisées (c'est le renouvellement cellulaire), souvent dédié à des cellules spécialisées (ou cellules souches) qui présentent la plupart du temps des divisions formatives. En bleu, les quantités d'ADN par cellule (2q étant la quantité unité dans une cellule diploïde) et nombre de chromosomes (2n étant le nombre de chromosomes dans une cellule diploïde); la phase S (synthèse) du cycle cellulaire est la phase de réplication ou duplication de l'ADN. La lettre G fait référence au mot anglais "gap" désignant un fossé ou état stable de la cellule (d'après la source citée, fig 1, p 498). Au cours du cycle cellulaire on différencie l'interphase (qui comprend les phases G1, S et G2) et la phase de division cellulaire (M, qui comprend deux phénomènes plus ou moins imbriqués: la mitose, ou division nucléaire, et la cytodiérèse, ou division cytoplasmique) qui correspond habituellement à moins de 10% de la durée du cycle cellulaire.
|
Tableau comparatif des divisions cellulaires eucaryotes et procaryotes |
||||
|
|
|
|
||
|
|
|
|||
|
matériel génétique |
duplication |
pendant la phase de croissance précédant la phase de division |
en interphase (phase S) avant la division (phase M) |
|
|
séparation |
par accrochage de chaque boucle d'ADN à la paroi en croissance (phase de division) |
par accrochage des chromosomes par les kinétochores de microtubules spécifiques (kinétochoriens) glissant le long des microtubules du fuseau (anaphase de la mitose) |
||
|
séparation des deux cellules filles |
élaboration d'une paroi intermédiaire |
étranglement cytoplasmique |
élaboration d'une paroi intermédiaire |
|
|
matériel responsable des mouvements |
pas de mouvements autres que la croissance : pas de microtubules ni de vésicules |
microtubules et filaments d'actine, vésicules golgiennes centrioles dans le centre organisateur des microtubules |
microtubules et filaments d'actine, vésicules golgiennes pas de centrioles dans le centre organisateur des microtubules mais une bande préprophasique puis un phragmoplaste au niveau de la zone de partage cellulaire |
|
|
moteur de la séparation (division bactérienne ou cytodiérèse des eucaryotes) |
allongement de la paroi, rupture de la cellule (synthèse de paroi et/ou déformation ????) |
microtubules du cytosquelette puis rupture de la cellule par étranglement |
microtubules du cytosquelette pour tous les organites y compris les chromosomes; la paroi ne venant séparer que tardivement deux compartiments; pas de rupture de la cellule mère mais un partage de l'espace |
|
Sources de cette partie:
Physiologie végétale, R. Heller (avec R. Esnault et C.
Lance), Masson, 1994; un incontournable
Biologie végétale, tome 2 : Organisation
végétative, D. Robert et A.M. Catesson, Doin,
1990
Le développement regroupe l'ensemble des événements qui, chez les pluricellulaires (plantes et animaux), vont du zygote à l'organisme complet et mature. Le zygote est la cellule reproductrice fécondée. C'est le point de départ quasi général du développement, même s'il est entouré de nombreuses cellules ou n'est pas même unique, comme chez les plantes à fleurs. On peut donc inclure dans le développement les étapes nécessaires à l'acquisition de l'état reproducteur. Le développement comprend la morphogenèse qui désigne étymologiquement les étapes qui conduisent à l'apparition et au maintien de la forme (du grec morphos = forme), non seulement de l'organisme, mais encore des organes que l'on désigne spécifiquement sous le terme d'organogenèse.
La mérèse (terme
qui vient du grec "merein" = partage)
désigne la division cellulaire chez
les plantes.
Le pouvoir de se diviser est assez général pour toutes
les cellules embryonnaires, comme dans l'embryon animal. Dès
que les premiers tissus se forment on peut repérer des zones
spécialisées dans la division cellulaire: ce sont les
méristèmes.
Les méristèmes primaires sont à la fois
histogènes, c'est-à-dire qu'ils forment les
tissus ("histo":= tissu), ensemble de cellules
spécialisées (différenciées)
réalisant une fonction commune, et organogènes :
ils forment les organes que sont les racines et la tige
ramifiée et feuillée et les fleurs. Ils assurent la
croissance vers le haut et le bas de la plante.
Les méristèmes secondaires, strictement
histogènes, mettent en place des tissus secondaires
protecteurs (liège) et conducteurs (bois...). Ils
n'existent que chez les Ptéridophytes (fougères) et les
Angiospermes dicotylédones mais sont responsables de la
croissance en diamètre de nombreuses plantes de ce groupe qui
ont un port arborescent (nombreux arbres).
La persistance des méristèmes tout au long de la vie de la plante implique que le développement n'est jamais terminé (on parle d'embryogénie indéfinie). La feuille, qui ne possède pas à proprement parler de méristèmes (les zones de divisions sont dispersées) a une embryogénie limitée dans le temps.
Cependant, les cellules de plantes, même différenciées et normalement incapables de se diviser peuvent retrouver cette capacité lorsqu'elles sont isolées. C'est ce qui autorise d'innombrables cas de reproduction asexuée naturelle ou artificielle, notamment par des cultures in vitro. La capacité d'une cellule isolée (ou d'un fragment) à redonner un organisme pluricellulaire complexe entier (capacité organogène et histogène) est qualifié de totipotence. Un organisme issu d'une seule cellule (prélevée sur un organisme adulte et non pas résultant d'une fécondation) et possédant une information génétique identique à cette cellule mère (et donc à un autre organisme) est qualifié de clone. Deux clones sont deux organismes génétiquement identiques (ce terme est en fait assez vague et peut prêter à bien des confusions, voir page sur les manipulations cellulaires).
Comment la mérèse est-elle contrôlée
chez les plantes ?
Un développement embryonnaire est avant tout autonome.
La capacité à se diviser et la totipotence sont des
propriétés partagées par toutes les cellules
embryonnaires, puis par des cellules spécialisées. Mais
cette capacité n'est pas perdue définitivement et peut
être réactivée pour de nombreuses cellules.
Cependant, l'obtention d'un végétal complet à
partir de cellules banales différenciées ou de cellules
sexuelles isolées non fécondées
(parthénogenèse) restent des manipulations difficiles
et il est bien plus facile d'obtenir un nouveau plant par culture
in vitro de méristèmes.
On notera que la totipotence ne s'exprime qu'avec la mise en place
d'une colonie de cellules (clone qualifié de
"cal", Belin p 125) à partir de laquelle se
différencient des organes (organogenèse) qui
donneront toutes les parties de la plante qui peut atteindre la
maturité sexuelle. Cependant, si l'on procède à
des repiquages successifs, cette compétence
organogène, capacité à mettre en place une
organogenèse à partir d'une colonie de cellules, se
perd. On obtient bien alors des colonies de cellules mais elles sont
perdu la capacité à former une plante. Ce que l'on peut
comparer à ce que donnent les cellules animales en culture: en
effet la plupart des cellules prélevées chez l'adulte
peuvent tout au plus reformer un tissu en culture mais sont
incapables de donner un individu entier. Certaines cellules souches
adultes sont cependant capables de donner plusieurs types cellulaires
mais il n'en reste pas moins que seules les cellules embryonnaires
animales possèdent une totipotence (pour
quelques bases à ce sujet voir page
sur les cellules souches).
Lorsqu'on isole une cellule végétale on rompt le lien cytoplasmique qui unit les cellules entre elles par les plasmodesmes. C'est donc une technique induisant un traumatisme. On peut aussi favoriser la reprise des divisions des cellules isolées en leur ôtant leur paroi en réalisant ce qu'on appelle un protoplaste ou cellule végétale sans paroi.
TP - mérèse et méristèmes
L'auxèse (de "auxi" parfait du verbe latin "augere" = "faire croître" ou du grec "auxè" = "la croissance") désigne l'extension de la cellule végétale. C'est principalement une élongation, c'est-à-dire une croissance en longueur (dans le sens d'allongement des organes de la plante: racine ou tige feuillée).
On pense que cette extension est due à un relâchement de la paroi intervenant simultanément avec une poussée de turgescence. Cette extension est sous le contrôle de régulateurs chimiques de croissance.
La constitution de la paroi et son extensibilité
La paroi n'est extensible que pendant une
brève période de croissance cellulaire qui fait suite
à la cytodiérèse et cesse en tout cas
après la différenciation.
Le phragmoplaste mis en place à la fin de la
cytodiérèse va donner une zone intermédiaire
entre les deux cellules végétales nommée
lamelle moyenne. De part et d'autre de cette lamelle vont
être apposées des composés glucidiques
(cellulose, hémicellulose, pectines..) et peptidiques
(extensine...) qui vont former une paroi. On distingue deux
étapes:
* une paroi primaire, mince et relativement souple (en tout
cas extensible) à l'origine, composée de couches
intercalées de cellulose (principalement) et d'autres
éléments formant une matrice.
* une paroi secondaire, très épaisse,
inextensible, composée de couches régulières de
cellulose; elle n'apparaît que chez quelques cellules.
La paroi peut subir de nombreuses modifications postérieures
à son dépôt. Par exemple, elle peut
s'imprégner de composés divers (subérine,
lignine, minéraux...) qui la rigidifient et empêchent
son extension.
Remarque:
La paroi n'est bien sûr pas inerte et elle n'a pas qu'un
rôle de cytosquelette externe. L'ensemble des parois, lamelles
moyennes et méats de la plante forme ce que l'on appelle
l'apoplasme que l'on appose au symplasme formé
par la réunion de tous les cytoplasmes des cellules en
communication les uns avec les autres par l'intermédiaire des
plasmodesmes. Il ne faut pas oublier que la membrane plasmique de la
cellule végétale est l'objet d'un intense remaniement
comme toutes les structures cellulaires. On estime que chaque minute
10% de la surface totale de la cellule sont renouvellés, ce
qui fait un renouvellement total en 10 min
(Robert et Roland, p
64). Les vésicules du REL et du Golgi
qui contiennent les éléments pércurseurs de la
paroi participent de cet intense traffic.
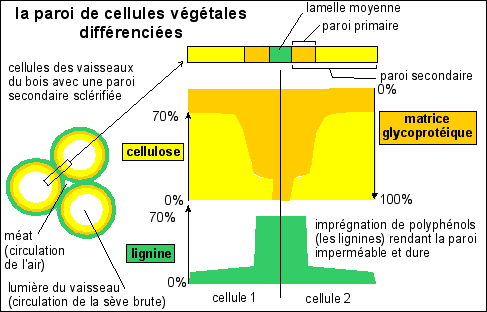
La cellulose est un glucide (sucre). C'est un polymère de ß-glucose. C'est une molécule orientée qui s'assemble en microfibrilles ordonnées. La cellulose est le composé organique le plus abondant sur terre dont la production est estimé à 50 à 100 Gt.an-1, ce qui correspond à la moitié de la biomasse terrestre. L'homme ne digère pas la cellulose, par manque d'équipement enzymatique adéquat (des cellulases), et la cellulose n'a donc aucune valeur nutritive pour lui. Les ruminants (vache ...) ne sécrètent pas non plus de cellulases mais ils hébergent des bactéries et des unicellulaires synthétisant des cellulases et libérant du glucose pour le mettre partiellement à disposition de l'animal qui consomme aussi les bactéries et les unicellulaires lors de la rumination.
L'extension est un phénomène irréversible.
Elle ne se passe pas dans le domaine élastique mais dans le
domaine plastique (pour des précisions sur le
vocabulaire concernant la déformation, voir cours
général sur les
séismes).
Elle comporte une composante élastique mais aussi une
composante dite plastique qui correspond à cette
irréversibilité de la croissance.
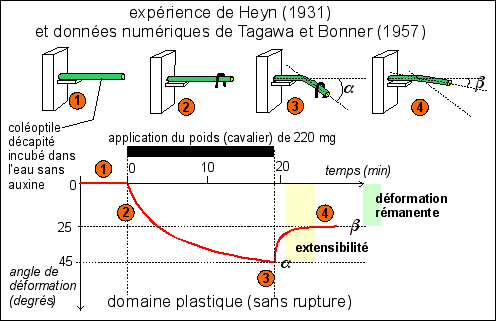
On peut mettre en évidence la plasticité au niveau d'un
organe en mesurant la courbure d'une toute jeune tige de
graminée (le coléoptile forme une gaine qui entoure la
première feuille; il croît par division de ses cellules
terminales latérales qui forment un anneau
méristématique et par allongement des cellules issues
de ces divisions - voir TP). On mesure alors la plasticité des
cellules en cours d'allongement, déjà
éloignées du méristème terminal qui a
été sectionné dans ces expériences
("décapitation" du coléoptile).
Il faut noter la forte cohérence du réseau de microfibrilles de cellulose enchâssées dans une matrice elle-même très structurée (à la façon d'un contreplaqué composé de plusieurs feuillets successifs dont les feuillets de fibres correspondraient à la cellulose et la colle à la matrice). L'extension s'oppose à cette structuration. On pense que l'extension serait due avant tout un relâchement du réseau des microfibrilles de cellulose par déstabilisation de la matrice. Mais les théories sont encore très discutées. Il est hors de question de s'intéresser aux mécanismes moléculaires à ce niveau d'enseignement.
L'auxine (AIA: acide indole-3-acétique) est la première substance chimique que les phytophysiologistes s'accordent à nommer hormone végétale car elle est synthétisée dans les méristèmes, transportée (de façon assez nettement polarisée dans le sens vertical descendant mais on ignore son mécanisme et sa voie précise de transport) et agit in vivo loin de son lieu de production (elle favorise l'histogenèse, l'organogenèse des racines et l'élongation des tiges et du coléoptiles des Graminées).
Expériences à analyser:
* Darwin (1880); Bordas A2 p 118 et Nathan p 133
* Boysen-Jensen (1910-1913) - Bordas p 119 B3 et Nathan p133
* Paàl (1919) - Bordas p 119 B4 et Nathan p133
* Söding (1923-1925) - Bordas p 119 B5
* Went (1928), Thimann (1934) - Bordas p 120 et Nathan p133
Mais nos connaissances sont extrêmement fragmentaires. Les rôles des auxines et autres médiateurs sont souvent paradoxaux selon les dosages et antagonistes sur différents organes. Actuellement, je ne pense pas qu'il y ait encore une vue d'ensemble sur ces médiateurs.
|
|
10-8 (0,05) |
10-7 à 10-6 (0,5 à 5) |
10-5 (50) |
|
|
élongation des organes |
coléoptile, tige, pétiole, limbe (Monocotylédones) |
|
|
|
|
limbe (sauf Monocotylédones) |
|
|
|
|
|
racine |
|
|
|
|
|
prolifération cellulaire |
cambiums (méristèmes secondaires) et tissus de réserve des fruits |
|
|
|
|
zone d'abscission (rupture du pétiole ou pédoncule floral avec la tige permettant la chute des feuilles ou des fruits) |
|
|
|
|
|
histogenèse |
|
|
|
|
|
organogenèse |
organogenèse de bourgeons (apparition de bourgeons donnant des tiges feuillées) |
|
|
|
|
organogenèse de racines (rhizogenèse) |
|
|
|
|
On ne connaît pas avec certitude les mécanismes moléculaires de l'action de l'auxine. Une action directe (ou indirecte) sur l'expression de l'information génétique est fortement suspectée (tant au niveau de la transcription que de la traduction).
De très nombreux
régulateurs de croissance ont été
isolés. On peut citer les gibbérellines (famille
des terpènes, agissant sur l'allongement des
entre-nœuds), les cytokinines
(dérivés de bases puriques agissant sur la
multiplication des cellules de la moelle de Tabac - test de
Skoog) et l'éthylène
(CH2 = CH2, agissant sur la maturation
des fruits qu'il accélère).
Toutes ces substances agissent en synergie (ensemble); leur
production et leurs rôles sont variées selon les organes
et les conditions environnementales (température,
éclairement...).
TP - auxèse
* (voir page spéciale) Des caractères des individus et des espèces
* Remarques sur cette partie du programme
La vision du vivant que votre programme
affiche en avant propos de cette partie est franchement
dépassée (l'artifice unificateur du
phénotype-génotype pour toutes les parties du programme
de 1èreS est particulièrement difficile à
justifier, même si un des souhaits affichés (dans les
documents d'accompagnement) est celui de casser une vision trop
étroite de la liaison génotype-phénotype) : Voici une formulation des
mêmes notions, à mon avis moins
obsolète Classifications des espèces Les êtres vivants sont classés en
espèces qui se ressemblent et dérivent les
unes des autres. La morphogenèse
végétale et l'établissement du
phénotype La morphogenèse
végétale « Le phénotype morphologique d'un individu
est le résultat des interactions entre l'expression
du génotype et son contrôle par
l'environnement. L'établissement de ce
phénotype met en jeu un ensemble de processus
biologiques dont des gènes sont responsables (mitose,
métabolisme cellulaire, action d'hormones, mise en
place des structures de l'organisme). Les
gènes gouvernent à la fois les grands traits
de l'organisation et les détails de la
structure, en permettant la synthèse de
protéines spécifiques aux diverses
échelles qui constituent l'organisme (cellules,
tissus, organes, plan d'organisation).
L'expression de ces gènes est soumise
à des facteurs externes (abiotiques ou biotiques)
dont la variabilité s'ajoutent à la
diversité allélique pour aboutir à une
diversité phénotypique
individuelle. La forme d'une plante adulte est déterminée
par des facteurs internes et externes pouvant jouer à
toutes les étapes de son développement qui
peut durer des années et même des
siècles. Quelques explications: Cependant... Nos connaissances sur la morphogenèse, la
genèse de la forme donc, sont pour le moins
incertaines, jamais réductibles à
l'échelle moléculaire, et très
théoriques, ce qui rend quasiment impossible leur
enseignement à ce niveau. Traiter des
méristèmes ou de la phyllotaxie nous passionne
tous mais pas dans le cadre d'un "phénotype" et pas
avec 6 heures de cours....
Le programme utilise le mot
espèce plusieurs fois il est nécessaire de le
définir.
L'étude de la morphogenèse des
végétaux permet d'aborder dans un cadre
intégré ces différents
phénomènes qui contribuent à
l'établissement du phénotype ».
- Les facteurs internes à l'organisme sont en partie
hérités, d'une façon stable au sein de
l'espèce. L'information génétique en
est une partie mais il faut sans aucun doute lui ajouter une
information cytoplasmique sans laquelle aucune transmission
de l'espèce n'est possible. Le travail du
développement est fondamentalement autonome. La part
des informations génétiques, cytoplasmiques et
environnementales n'est pas la même à toutes
les étapes du développement: les informations
génétiques et cytoplasmiques semblent
primordiales dans les premières étapes du
développement. Une fois l'organogenèse
commencée les différentes populations
cellulaires échangent des informations de type
environnemental, comme les hormones et autres messagers
chimiques, qui permettent un fonctionnement
intégré de l'organisme vivant. Une
caractéristique essentielle du développement
réside dans le mémoire que les cellules
embryonnaires ont des étapes antérieures
qu'elles ont suivi, même si les influences auxquelles
elles avaient été soumises alors ont
cessé (voir Biologie
moléculaire de la cellule, Alberts et al., 1994
(3ème éd.), p 32 eu page sur une
théorie
du
développement).
- Les facteurs externes à l'organisme, ou
environnementaux, jouent de façon importante au cours
des premières étapes du développement
puis peuvent orienter tel ou tel aspect du
développement conduisant à des formes
similaires chez des espèces différentes
(convergence) et des formes différentes au sein de la
même espèce (variabilité
intraspécifique).
À mon avis, il serait plus judicieux encore de
replacer correctement dans leur contexte scientifique (et
non plus scolaire) les notions de génotype et de
phénotype et ainsi les problèmes se
résoudraient tout seuls. Quelques points à
soulever à mon sens :
* Un gène est une unité de fonction
strictement moléculaire : c'est une information
pour une molécule. Or la vie est un
phénomène cellulaire que personne ne peut
réduire à des molécules, si ce n'est
par idéologie. La cellule reste le niveau
irréductible de compréhension du vivant. Voir
la page sur les niveaux
d'organisation du vivant, le cours de
seconde sur la cellule, et la page sur la
biodiversité qui présente aussi une
théorie du
vivant.
* Ainsi, pour un pluricellulaire, le rôle des
gènes -qui ne peut être défini qu'au
niveau de chaque cellule- ne peut être
extrapolé de façon simple à l'individu
entier. Le génotype et le phénotype sont
des notions cellulaires. Voir page sur
les génotypes et les phénotypes.
* La biologie de l'individu dépasse le niveau
moléculaire auquel se cantonne la
génétique, même si elle
s'intéresse à ces molécules aux niveau
des populations. Les lois de l'organisation du vivant
sont décrites par des
théories de la forme et de
l'évolution dont nous reportons
l'étude succincte à la classe de terminale
lorsque nous étudierons les caractères. Voir
cours de terminale sur la
biodiversité et l'histoire des êtres
vivants.
L'intitulé du chapitre: "La morphogenèse
végétale et l'établissement du
phénotype" est exagérément ambitieux et
je crois qu'il est impossible de traiter la
morphogenèse végétale en 1ère S
avec le volume d'heures imparti. D'où mon plan,
provisoire (mérèse et
auxèse).
Une fois encore la notion de phénotype (au sens de la
biologie moléculaire) n'est pas
généralisable à un individu complet.
Cette généralisation est à mon avis
encore plus inadéquate chez individu pluricellulaire
dont le cytoplasme de toutes les cellules est plus ou moins
commun (symplasme) et dont l'apoplasme constitue une voie
essentielle de communication (sauf entre la plante
mère et l'embryon par exemple) et dont la totipotence
cellulaire est reconnue pour de nombreuses cellules
différenciées...pour ne citer que deux
propriétés particulièrement
frappantes.
L'embryogenèse végétale se
différencie de l'embryogenèse animale par deux
caractères principaux:
- elle s'effectue sans migration cellulaire; on
retrouve ici un point capital de l'analyse du
développement si cher aux défenseurs de la
génétique du développement qui
travaillent à partir de l'hypothèse d'un
programme génétique; la place des cellules
étant fixée (ici de
façon permanente par leur paroi commune, alors que
chez la drosophile la place est fixée de façon
"dynamique", dans le sens de "non expérimentale",
dans les enveloppes de la pupe que l'on ne peut ouvrir pour
effectuer des expériences d'embryologie
expérimentale - voir page
sur le
développement)
la plupart des modifications génomiques induisent des
modifications morphologiques reproductibles. Ce qui
n'est très généralement pas le cas si
l'on déplace les cellules par embryologie
expérimentale notamment.
- elle s'effectue sans connexions vasculaires entre
la plante-mère et l'embryon: l'embryon est soit libre
dans un milieu aquatique, soit entouré d'une paroi
dépourvue de plasmodesmes (connections cytoplasmiques
entre cellules adjacentes); les échanges de
substances entre la plante-mère ou le milieu
extérieur et l'embryon se font donc par voie
apoplasmique (l'apoplasme est le milieu formé
par les parois des cellules et les espaces situés
entre les parois (méats) où circulent des
solutions aqueuses et de l'air; il s'oppose au
symplasme qui représente l'ensemble des
cytoplasmes cellulaires réunis par des
plasmodesmes).