|
|
||||||||||||||||||
|
Plan |
|
|
|
|||||||||||||||
|
1 - Héritage thomien Depuis que
j'ai
goûté
à
René
Thom, je n'ai
pu me
défaire
d'une
formalisation
attirante
même si
elle demande
un effort
intellectuel
vers les
mathématiques
que je suis
loin d'avoir
accompli. Thom
n'a pas non
plus vraiment
facilité
la tâche
aux
biologistes en
mélangeant
les allusions
philosophiques,
mathématiques,
physiques et
biologiques au
gré de
sa culture. Je
tiens Notre propos est l'hydratation. Je comprends ce thème comme la place de l'eau dans la vie de l'organisme humain et tout particulièrement dans la fonction de nutrition. On s'accorde en France à faire reposer la vie sur trois grandes fonctions globales non localisables que je me plais à qualifier de travail : le travail de nutrition, souvent exprimé sous forme d'échange de matière et d'énergie, le travail de reproduction, qui est fondamentalement une fonction de l'espèce plutôt que de l'individu et enfin le travail de relation, qui correspondent à l'expression des échanges d'information, que l'on pourrait qualifier d'aspect social de la vie.
|
|
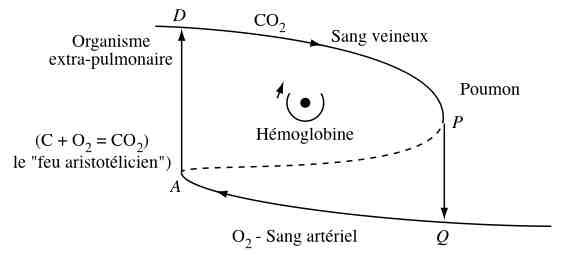
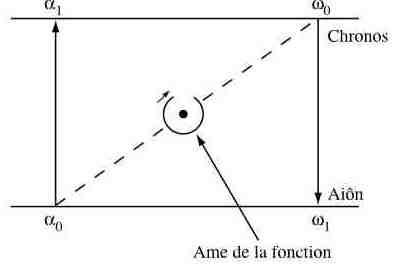 in Structure et fonction en biologie aristotélicienne, 1988f7, fig5 Pour René Thom, toute fonction est la manifestation (ci-dessus) d'un pli des temps (aiôn en grec, le temps intemporel où il ne se passe rien, ce qui correspond aux variables lentes d'un système) sur l'espace-temps (chronos, porteur d'événements catastrophiques, ce qui correspond aux variables rapides du système). La dynamique cyclique correspond à la catastrophe fronce (une des 7 catastrophes élémentaires qui géométrisent les dynamiques). La flèche verticale α0->α1 correspond au début de l'action, l'excitation, apparition d'une forme spécifique pour le système; la flèche verticale ω0->ω1 correspondant au retour au repos. Le centre est "l'âme" de la fonction, le point organisateur, qui ne correspond pas forcément à une structure. |
||||||||||||||||
|
Les banalités habituelles plus ou moins exactes -eau milieu de vie , eau indispensable à la vie, eau solvant du vivant , ... - prennent un sens vraiment signifiant et peuvent être corrigées dans le schéma fondamental de René Thom qu'il appelle la blastula physiologique qui est issue d'une double duplication du cycle physiologique (nutritif) fondamental. Dans le cycle du haut, on notera que pour respecter la position embryologique l'aiôn du pli des temps est vers le haut (côté dorsal embryonnaire) alors que le chronos du pli des temps est en bas (côté ventral embryonnaire). C'est l'inverse pour le cycle du bas. 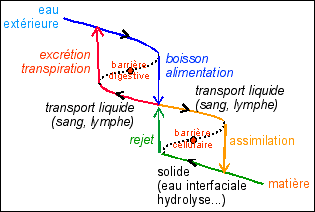 Une représentation de la dynamique de l'eau dans l'organisme à la façon de René Thom. Les choix faits ici des "âmes" de chaque fonction sont évidemment discutables. Je n'ai pas non plus distingué les changements de phase de l'eau liquide <-> gaz, ce qui obligerait à rajouter un cycle respiratoire au cycle du haut. Il est possible aussi qu'au niveau des cellules des changements d'état de même type interviennent ce qui obligerait aussi à dupliquer le cycle du bas. (Enfin, le problème majeur pourrait être qu'il faille en fait considérer une seule âme à la fonction dupliquée et donc changer la représentation du cycle sur une surface unique en deux surfaces : catastrophe papillon par exemple ? Ce qui dépasse mon niveau de compréhension). Nous nous intéresserons aux étapes d'assimilation / rejet en utilisant le formalisme de Thom qui permet de relier intelligemment structure-fonction (voir extrait ci-contre) : la fonction de l'organe (ou de la structure, tissulaire, cellulaire...) détermine sa forme, c'est une expression du lien entre les différents types de causalité (et particulièrement finale et formelle). Le fait que l'on retrouve la catastrophe fronce, dont les caractéristiques seront développées plus bas, de façon omniprésente est la marque de la cyclicité des phénomènes. |
|
René
Thom, Structure
et fonction en
biologie
aristotélicienne,
1988f7,
pp13-14 «
Chez les
vertébrés
(supérieurs)
le centre
organisateur,
l'âme de
la fonction
«
circulation
», s'est
localisé
dans le
cœur ;
il s'est
réalisé
spatialement
par
l'accolement
de deux tubes
pulsatiles
correspondant
aux «
links »
des
arêtes
DP, QA de la
figure 8
[ci-dessous]
plongée
dans R3
. Je
proposerai
pour la
représentation
formelle de la
relation
structure-fonction
les principes
suivants : |
||||||||||||||||
|
2 - "Deviens ce que tu manges" Nous allons nous intéresser à la partie haute du double cycle : alimentation-boisson. |
||||||||||||||||||
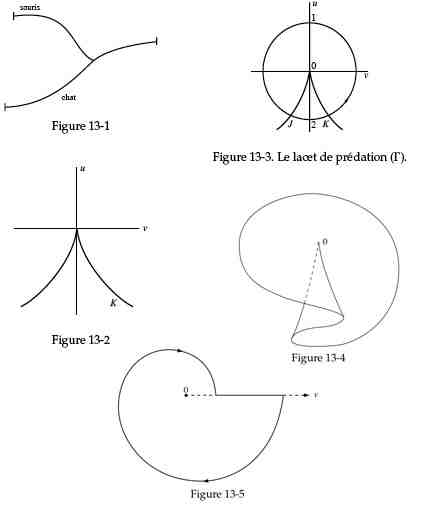 Le lacet de prédation par René Thom (figures extraites de Stabilité Structurelle et Morphogénèse : essai d'une théorie générale des modèles, 1967); Fig 13-1 : le chat mange la souris : catastrophe de capture, Fig 13-2 et 13-3 : projection de la fronce dans le plan (u, v); Fig13-4 : surface F de la fronce de capture ; Fig 13-5 : projection du lacet de prédation dans le plan (a, b) ou (u,v). (Je recommande aussi la représentation de P. Lacorre, Sur un nouveau type de représentation catastrophiste pour les modélisations en biologie et sciences cognitives, Intellectica, 24, 1997/1) |
|
Pour l'homme, se nourrir c'est assimiler les autres. L'homme se nourrit "des autres". On dit qu'il est "allotrophe" - la racine allo = autres, est à mon avis préférable à hétéro = qui signifie plutôt différent. Se nourrissant d'autres êtres vivants -vivants ou morts - c'est un prédateur au sens où toute nourriture devient une proie à capturer et à assimiler par la digestion. René Thom affirme "le prédateur affamé est sa proie"; voir la notion de lacet de prédation (voir illustration ci-contre) dans l'œuvre de Thom; voir aussi Les chemins du sens à travers les sciences, AL, [1984, 9. 3] (1984f9.pdf); auquel on peut aussi ajouter le travail de Philippe LACORRE, Sur un nouveau type de représentation catastrophiste pour les modélisations en biologie et sciences cognitive, Intellectica, 1997/1, 24, pp. 109-140). La nutrition allotrophe est une catastrophe de capture: deux actants, le prédateur et la proie, n'en font plus qu'un, à la fin de l'action. Il y a ici le cycle fondamental d'un retour à un niveau stable, antérieur à la capture et à l'assimilation (1967f8.pdf, p3s). La
nutrition est
la
satisfaction
d'un besoin.
Un manque de
nourriture ou
d'eau est une
instabilité.
La fonction de
nutrition
globale peut
être
approchée,
localement
(par exemple
au niveau
d'une
cellule), par
un potentiel
et l'acte de
se
nourrir-boire
par le retour
du
système
à une
valeur
minimale du
potentiel. La
nutrition est
toujours
accomplie
lorsque
l'organisme
minimise son
potentiel. La stabilité est une des caractéristiques des dynamiques du vivant. Ce n'est pas le système vivant qui est stable (ou immobile) mais sa dynamique, c'est-à-dire ses fonctions. On dit encore que l'organisme est en équilibre dynamique (qu'il fonctionne), ce qui est le propre d'un système en homéostase. Ce terme vient du grec homéo (ou homo = identique) et stase qui signifie position. La propriété d'un système en homéostase est l'homéostasie. Un système homéostatique est un système qui est stable face à de petites perturbations. Afin de déterminer le déploiement d'une fonction dans l'espace de régulation il faut s'intéresser à la topologie de la surface de catastrophe, c'est-à-dire le lieu où "il se passe quelque chose". En chaque point de cette surface la dynamique est sous-tendue, dans la théorie des catastrophes, par un conflit entre attracteurs. Il faut noter cependant que l'on ne peut pas connaître la morphologie de toute la surface étant donné la complexité du vivant et l'indéterminisme associé (certains attracteurs étranges peuvent ne pas être formalisables). On se contente donc de quelques cas simples où la dynamique peut être formalisée. |
||||||||||||||||
|
Dans la catastrophe de nutrition, la partie digestive correspond aux grandes valeurs de u (partie à variation lente où il n'existe qu'un seul et unique attracteur). La digestion est une assimilation de la proie par le prédateur. C'est le devenir de toute nourriture : participer à la matière et à l'énergie du prédateur-consommateur. En cela l'eau de boisson ou de perfusion ne peut pas différer des autres substances nutritives solubles ou non, y compris les gaz. |
|
Ce n'est que dans le cas où l'on séparerait le rôle de l'eau des autres substances que l'on pourrait imaginer des fonctions différentes. L'eau serait en quelque sorte inerte, sans pouvoir nutritif. Mais c'est justement l'idée inverse qui ressort des études cytologiques. Les rôles de l'eau, tant structuraux que fonctionnels, en font un élément essentiel du métabolisme qui est capable de fournir de la matière bien évidemment, mais aussi de l'énergie, non pas par une dégradation où une dissociation (comme dans le cas de la photolyse), mais par ses liaisons aux autres molécules. L'énergie de liaison apportée par l'eau est peut-être l'énergie principale des événements cellulaires. Non pas comme un solvant, mais bien comme un élément structural, de communication...etc. (revues dans P. Mentré et aussi un peu ici) Donc l'eau
nourrit
l'organisme
et participe
de la
matière
et de
l'énergie
du
consommateur
au sein de la
fonction de
nutrition. |
||||||||||||||||
|
|
||||||||||||||||||
|
3 - L'interface membranaire lieu de catastrophe |
|
|
||||||||||||||||
|
|
|
3.1 - Données cytologiques Avant de pouvoir formaliser le rôle de la membrane cellulaire -au sens large d'interface - que j'ai pris comme âme de la fonction d'assimilation/rejet, je me suis efforcé de clarifier quelques données cytologiques en traduisant notamment deux articles - que je mets à disposition (sans autorisation) sur ce site Porter 1984 et Penman 1995 - cités par P. Mentré dans son diaporama auquel je renvoie . |
||||||||||||||||
|
Dans une vision moléculaire statistique, le chimiste voit la matière vivante comme un cas particulier de son objet d'étude qu'il s'efforce de comprendre à l'aide de lois universelles (ses théories) sous le contrôle de l'expérience. Le vivant est pour lui un milieu aqueux qui diffère du milieu artificiel par la grande variété de ses composants (à faible concentration). Sa compréhension de la compartimentation spatiale et temporelle est fortement influencée par les résultats des techniques d'étude employées. La microscopie électronique a conduit à surestimer le rôle des membranes (lipophiles et osmiophiles) (voir Penman 1995 qui n'hésite pas à proposer un nouveau modèle de structure cellulaire basé sur des lamina protéiques (externe et nucléaire) sur laquelle viennent s'insérer d'une part les bicouches phospholipidiques qui servent de filtre aux protéines solubles et d'autre part les réseaux du cytosquelette pris au sens large contenant non seulement les habituels filaments (microfilaments, filaments intermédiaires et microtubules qui composeraient 10-15% de la matrice) mais bien d'autres types qualifiés autre part de microtrabécules (Porter 1984) ou de "core filaments" (Penman 1995) dans le noyau...). En effet la théorie membranaire domine toute la compréhension des mouvements de matière entre la cellule et le milieu extérieur. De la même manière, l'étude des solutions aqueuses a conduit à favoriser la formulation des dynamiques en terme d'équilibres - ou de déséquilibres - dans des conditions isothermes et isobares avec des grandeurs thermodynamiques comme l'enthalpie libre. Les biophysiciens savent pertinemment que ces modèles ne reflètent que très mal la réalité, mais les efforts en direction de la théorisation des forces moléculaires et ioniques spécifiques au vivant ou du moins explorées dans les systèmes vivants (je pense notamment aux travaux de Barry Ninham) restent très difficiles d'accès. |
|
Les travaux des cytologistes qui ont perfectionné les techniques de microscopie électronique afin de voir la cytomatrice (cytomatrix) (p.ex. Porter 1984, Penman 1995) n'ont pas non plus débouché immédiatement sur une compréhension meilleure de la cellule puisqu'ils ont dans un premier temps majoritairement considéré que les réseaux filamenteux découverts structuraient la cellule comme une armature (ce qui déplaçait le problème de la structure à d'autres molécules que les protéines, mais sans changer de paradigme). Enfin, il n'est pas rare que le raisonnement des biologistes se raccroche à la théorie de l'information génétique en supposant qu'une information inconnue existerait pour gouverner la synthèse et l'organisation de ces nouvelles structures non protéiques. C'est à nouveau oublier la richesse des concepts physiques pouvant aider à comprendre la genèse et la dynamique des formes structurales sans le recours à l'idée plus ou moins magique d'information. Les efforts des biologistes pour utiliser les outils de la physique et de la chimie en respectant l'originalité de l'objet vivant restent isolés (Mentré 2011). Par exemple, les expériences réalisées sous très haute pression dans le but de simuler les conditions intracytoplasmiques (voir P. Mentré et G. Hui Bon Hoa, 2001 ; Mentré, P., Hui Bon Hoa, G.: The effects of high hydrostatic pressures on living cells: a consequence of the properties of macomolecules and macromolecule-associated water. Int. Rev. Cytol. 201, 1-84 (2001), ont permis de mieux comprendre combien l'eau interfaciale était une clef de la dynamique des molécules et surtout des assemblages moléculaires. Il faut donc à la fois considérer les avancées sur les composants cellulaires (cytomatrix) mais aussi sur les rôles de l'eau (dont le rôle structural une fois compris ne conduit pas implicitement aux mêmes erreurs que pour des structures filamenteuses) en s'efforçant à une explication physique, voire mathématique, quelque en soit le support physique. |
||||||||||||||||
|
|
|
3.2 - Le fonctionnement de la fronce de René Thom |
||||||||||||||||
|
Dans le but de modéliser le changement de phase assimilation/rejet, je pars sur l'idée exprimée par P. Mentré selon laquelle la déshydratation est un phénomène relativement irréversible qui est lié à l'histoire cellulaire, à un vieillissement, prématuré ou non, à une perte de la capacité à retenir son eau et non pas à la seule disponibilité en eau. |
|
Je précise encore une fois que je vais proposer un modèle plus que spéculatif et cela sera aux praticiens de dire s'il peut ou non les aider. Sa validation ne peut être théorique, mais uniquement pratique (du point de vue mathématique il me semble exact, mais le fait qu'il reflète la réalité n'est pas défini par la théorie). |
||||||||||||||||
|
Un flux entrant d'eau correspond à une dynamique de capture alors qu'un flux sortant correspond à une dynamique de rejet; ces deux dynamiques pourraient correspondent à une catastrophe unique de type fronce qui correspond à la bifurcation d'un attracteur unique (flux net nul ou de référence) en deux attracteurs (chaque minimum stable du potentiel standard est relié à un attracteur, voir ici et là pour le vocabulaire). La fronce est la catastrophe probablement la plus courante pour laquelle quelques applications en biologie ont déjà été présentées (voir page sur les modèles et article d'Ivar Ekeland) : cycle cardiaque, action enzymatique... |
||||||||||||||||||
|
Dans la formulation thomienne x désigne le flux, c'est la variable de sortie. Les paramètres de contrôle u, v et w du potentiel standard de la forme F(x) = x4 + u x2 + v x + w doivent donc de préférence refléter l'aspect continu de la nutrition (voir page CTmodel pou des applets et des graphiques). w est inopérant sur la dynamique (il ne modifie que la valeur du potentiel) et ne sera pas considéré.
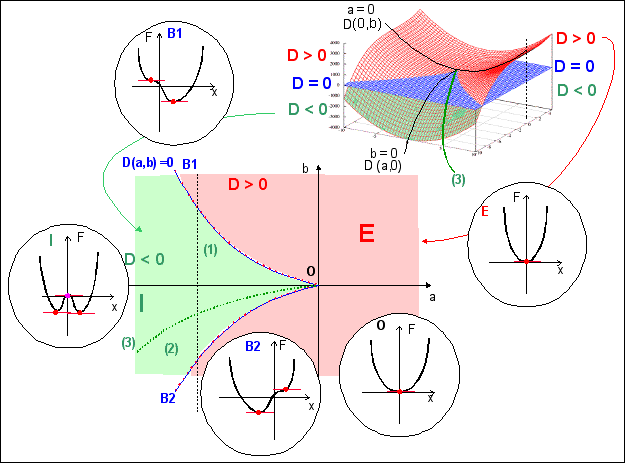 Ensemble de bifurcation de F (équation D(a,b)=0) et sections particulières de F(x,u,v) (cercles) où x prend des valeurs critiques (u =2a et v =4b) - voir page sur les modèles |
|
Pour étudier ce potentiel, on représente habituellement la surface des points singuliers (où critiques) de F où le potentiel présente un minimum stable (ensemble des points où dF/dx = 0). C'est une équation algébrique du troisième degré 4x3+2ux+v = 0 ou x3+ ax+b = 0 (avec a=u/2 et b=v/4) qui possède au moins une racine réelle et au plus trois racines réelles selon la valeur du discriminant D (D=4a3+27b2). Si D < 0, il y a trois racines réelles distinctes, pour D > 0, il n'y a qu'une racine réelle (et deux complexes conjuguées) et pour D = 0, il y a trois racines réelles, mais certaines coïncident (pour D = 0 et a Ç 0 ou b Ç 0, deux racines réelles sont égales et pour D = 0 et a = b = 0, les racines sont toutes trois égales). Le graphe correspondant à l'équation D(a,b)=0 dans le plan (a,b) est appelé "ensemble de bifurcation" de F. C'est l'ensemble des points (du plan (a,b)) où les "choses changent"; là où la stabilité de F "bifurque" en présentant un ou deux ou trois points de stabilité: ce sont des points de conflit entre régimes stables reliés mathématiquement à des attracteurs (chaque minimum stable est relié à un attracteur). L'ensemble des points de catastrophe est défini qualitativement par l'ensemble des points de conflit entre attracteurs. C'est sa forme qui donne le nom à la catastrophe, ici la fronce. 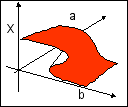
Une fronce (cusp) dans l'espace (x, a, b) dessinée par l'ensemble des points de catastrophe de la catastrophe élémentaire associée au potentiel standard F = x4 + ux2 + vx |
||||||||||||||||
|
|
|
|
||||||||||||||||
|
Dans la théorie des catastrophes, le principal problème est celui du choix des paramètres qui est toujours arbitraire. Choisir un paramètre biochimique est toujours possible, mais ramène à une vision discontinue dont on cherche à s'éloigner. Pour l'eau quel paramètre choisir qui puisse rendre compte de trois états possibles de la cellule ? On a en
effet un
état
où le
flux
résultant
est nul (ou
à une
valeur de
référence)
qui doit
obligatoirement
correspondre
à des
valeurs les
plus grandes
de a (donc de
u car u=2a), a
qualifie donc
un état
nutritif
stable
où
l'eau
accessible
ainsi que
l'eau produite
métaboliquement
sont
compatibles
avec les
dynamiques
internes de la
cellule
(croissance,
métabolisme...).
u ou a
mesurerait
ainsi l'état
sain. Cela
cadre bien
avec
l'idée
de
départ
selon laquelle
l'état
de
vieillissement
de la cellule
est le
paramètre
principal.Si
l'on reprend
les mises au
point faites
par les
physiologistes
sur la
question de
l'hydratation
ce
paramètre
pourrait aussi
bien
représenter
l'hydratation
totale
(qui baisse
avec
l'âge
mais pas pour
la masse
maigre) ou la
sensation
de soif (qui
baisse aussi
avec
l'âge).
Pour ce qui
est des apports
d'eau
(soit
alimentaire
soit de
boisson) le
paramètre
ne respecte la
séparation
entre
alimentation
et
hydratation,
ce qui
était
la question
première
de la
réflexion
posée.
Dans le
modèle
choisi, ces
deux
paramètres
ne peuvent
qu'être
équivalents. |
|
Lorsque u (ou a) diminue, on passe a un régime indéterminé (mais pas instable - la non-détermination vient du chemin parcouru et donc de l'histoire de la cellule -, voir l'applet ci-dessous pour appréhender cette mémoire; la stabilité vient du minimum et elle est assez robuste : pour v minimal on peut toujours faire varier u, on ne sort plus du puits de potentiel). v ou b (car v=2b) est le splitting factor (facteur d'écartement), c'est lui qui fait basculer la morphologie de la fonction (donc l'hydratation) à un état soit d'hydratation excessive (flux entrant excessif), soit à un état d'hydratation insuffisante (flux sortant ou entrant, mais insuffisant). Je vais prendre dans un premier temps, pour définir ce paramètre, le comportement physique de la barrière phospholipidique membranaire, où le fonctionnement des aquaporines qui ont été postulées. Selon l'efficacité de la barrière, l'eau entrerait trop ou pas suffisamment, ce qui signifierait que les mouvements en absence de barrière seraient toujours dans le sens extérieur vers intérieur.
Voici un applet (modifiée) de Lucien Dujardin (http://perso.wanadoo.fr/ l.d.v.dujardin/index.html) qui présente un modèle qui obéit à la règle de Maxwell: c'est le minimum le plus bas qui est toujours choisi et il y a une incertitude lorsque deux minima sont égaux (cette incertitude n'est pas prise en compte dans le modèle qui choisit l'une des deux racines pour représenter la couleur (morphologie) de sortie). Les trois racines de x3+ ax+b = 0 sont représentées par des traits verticaux rouges et le minimum est représenté par un trait horizontal rouge. |
||||||||||||||||
Vous pouvez voir le code de l'applet: cata3mh.java |
||||||||||||||||||
|
Que faire
d'un tel
modèle
?
Reflète-t-il
la
réalité
? |
|
Si
l'on s'efforce
aussi
d'utiliser le
modèle
pour
répondre
à la
question de
l'hydratation
en fin de vie
on a deux
éléments
essentiels
(bien visibles
grâce
à
l'applet): Pour lancer des protocoles expérimentaux qui iraient dans le sens d'une validation du modèle, en sachant que les paramètres ne sont pas mesurables in vivo à ma connaissance, il faudrait se tourner vers les résultats de l'hydratation dans des cultures vieillissantes. Je ne sais pas s'il en existe. |
||||||||||||||||
|
|
|
Je reviens maintenant à la question initiale de l'âme de la fonction représentée par l'interface cellule/milieu extérieur et non pas par la barrière membranaire, même si l'on ajoute les glycoprotéines, une lamina et un glycolcalix. Le lieu de l'assimilation / rejet est celui du changement de phase liquide -> solide. Non pas au sens de changement d'état de l'eau liquide -> glace solide, mais au sens d'un changement de paramètre qui de rapide devient lent. L'eau n'est plus mobilisable. Elle peut d'ailleurs même être impliquée dans une liaison covalente après dissociation et donc ne plus exister en tant que molécule d'eau. La topologie de la surface de changement de phase n'est évidemment pas celle de la surface cellulaire. |
||||||||||||||||
|
Que signifie fournir de l'eau par perfusion artificielle ? |
|
Les tissus
solides (phase
solide) sont
considérés
comme baignant
dans une
lymphe
interstitielle
(phase
liquide) et en
contact
-à
travers des
parois
vasculaires-
avec des
liquides
canalisés
(lymphe
canalisée
et sang).
Augmenter
l'eau par
perfusion
consiste donc
à
augmenter le
volume
-localement au
point de
perfusion- en
supposant que
c'est le
volume global
de la phase
liquide qui va
augmenter et
se
répercuter
sur la phase
liquide
interstitielle,
puis par
diffusion, sur
la phase
solide.
L'imaginaire
des
biologistes
considère
donc que l'eau
des
différents
compartiments
est
échangeable
avec des
coefficients
variables de
diffusion pour
chaque
séparation.
Le
problème
est que le
tissu n'est
pas une phase
solide. De la
même
manière,
l'eau
cellulaire n'a
pas du tout
les
propriétés
d'une phase
liquide et la
diffusion ne
peut pas
rendre compte
de son passage
au sein de la
cellule. |
||||||||||||||||
|
Il faut donc préciser la notion de phase. La phase d'un système (au sens de la thermodynamique) est définie comme une partie homogène séparée des autres parties du système par une surface. Mais cette surface n'est pas définie dans notre espace euclidien à 3 dimensions, mais dans un espace ou espace des phases où le système est défini simplement par ses coordonnées (les variables dynamiques du système). Toute modélisation d'un changement de phase nécessite donc la recherche des coordonnées les plus pertinentes. Lorsque l'eau passe du milieu "eau libre" au milieu "eau interfaciale" elle subit un changement de phase. On pourrait considérer qu'au voisinage des points d'entrée de l'eau (aquaporines avec canal hydrophobe) et entre les mailles d'un réseau de microtrabécules (Porter, 1984, Penman 1995) il existe des zones où l'eau cellulaire est essentiellement liquide, mais ce n'est que repousser le problème à l'intérieur de la cellule. Il est évident que l'on ne peut prétendre connaître dans le détail la forme de cette surface d'échange qui pourrait être extrêmement complexe et surtout mouvante. |
|
Quel
paramètre
pourrait
rendre compte
de
l'état
de l'eau ? |
||||||||||||||||
|
|
||||||||||||||||||
|
Remarque: JG
Watterson
propose donc
que : «
Dans le
modèle
du domaine
onde-cluster,
la force qui
sous-tend
l'action des
nanomachines
est la
même
force qui
maintient les
protéines
dans leur
forme, mais
aussi la
même
force de
pression qui
déplace
la
matière
à
travers les
membranes.
Elle
dérive
d'interactions
faibles ,mais
dynamiques ,
les liaisons
hydrogène,
qui exercent
les limites de
leur influence
jusqu'à
l'échelle
du pixel de
pression.
Au-delà
de cette
limite, une
pression
globale se
dégage.
C'est une
force
dirigée
vers
l'intérieur
: une force
qui rend
compte de la
construction
des structures
plurimoléculaires
et qui
s'oppose
à la
dispersion
causée
par
l'agitation
thermique».
(JG Watterson,
1997, extrait
de la
conclusion; «
In the
wave-cluster-domain
model, the
force
underlying the
operation of
molecular
nanomachines
is the same
force that
holds proteins
in their
stable form
and is also
the same force
that moves
water across
membranes
against
pressure. It
derives from
weak but
dynamic
interactions,
the H-bonds,
which can
extend their
bonding action
over long
ranges set by
the size of
the pressure
pixel. Below
this spatial
limit a
unifying
tension
operates. It
is an
inward-directed
force: a force
that supports
the build-up
of
multimolecular
structures and
opposes the
randomization
caused by
thermal
motion.») |
|
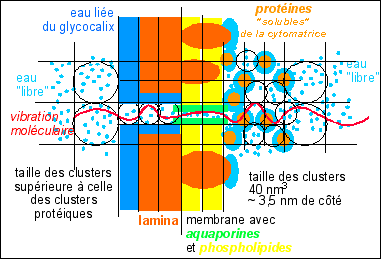 Le quadrillage de la cellule en clusters permet de comprendre la pression osmotique comme une grandeur reliée à la vibration des molécules au sein de chaque cluster. L'énergie des petits clusters, avec des protéines, est égale - à l'équilibre - à celle des plus grands clusters d'eau pure. La pression est donc supérieure du côté des clusters avec soluté. Les nœuds sont situés aus points où la vibration est minimale. Les cercles ont été ajoutés pour aider à donner une idée de l'échelle des domaines eau/protéines. Bein évidemment, seules quelques molécules d'eau sont représentées pour des clusters qui en contiennent environ 1300 pour les plus petits (inspiré de Watterson 1997, fig 3) - pour des tailles de molécules voir ici
Watterson trouve donc au sein de l'eau l'énergie nécessaire à rendre compte des l'organisation des clusters : les liaisons hydrogène. Les forces qui en découlent remplacent à cette échelle les notions de pression osmotique ou de potentiel hydrique... Cela n'empêche pas bien sûr des recherches plus approfondies sur les forces d'interactions à l'échelle du cluster qui sont loin d'être bien connues. Cette réponse est cependant un pont jeté entre l'échelle moléculaire discontinue et une fonction continue, et qui diffère de la réponse statistique habituelle. Elle diffère aussi de la voie thomienne qui s'intéresse non pas aux régularités, mais aux points catastrophes où il y a discontinuité. |
||||||||||||||||
|
La
théorie
des
modèles
de René
Thom permet de
modéliser
les points
où la
surface
présente
des
discontinuités,
des
rebroussements,
des points
particuliers.
Lors de la
nutrition,
l'assimilation
de l'eau par
les tissus et
les cellules
nous
paraît
être un
phénomène
régulier
sans
discontinuité,
sauf si
justement un
tissu ou une
cellule
"malade"
présentait
un
comportement
anormal. Un
excès
d'eau libre se
caractériserait
par une
diminution de
la surface de
changement de
phase alors
qu'un
déficit
en eau libre
pourrait
correspondre
à une
surface
agrandie,
bosselée,
voire
même
avec des
points de
rebroussement. |
|
La
quantité
d'eau libre
est donc
certainement
un
paramètre
important de
l'espace des
phases, mais
n'est-ce pas
justement
celui que l'on
cherche
à
atteindre ? Le
degré
d'hydrophilie
des composants
est ensuite un
paramètre
qui
paraît
incontournable
et qui semble
accessible
expérimentalement
par l'analyse
chimique. La
viscosité
est un
paramètre
qui n'est
envisageable
que dans une
phase fluide
unique, il est
donc
inapproprié
si l'on
considère
un changement
de phase. |
||||||||||||||||
|
|
|
|
||||||||||||||||
|
|
|
|
|
|||||||||||||||
|
|
|
J'ai
tenté
de justifier
le fait que
l'eau du
vivant
participait
d'une
finalité
nutritive et
que cette
finalité
était
à
l'origine de
formes
particulières
des
dynamiques,
mais aussi des
structures
cellulaires et
organiques. Je
suis bien
conscient de
la confusion
du discours,
qui utilise le
vocabulaire de
Thom sans
pouvoir tout
réexpliquer,
qui se
répète
souvent, et
qui
débouche
plus souvent
encore, soit
sur des choix
de
paramètres
franchement
arbitraires,
soit sur des
conclusions
qui sont des
évidences
quand elles ne
sont pas
incluses dans
les
hypothèses
de
départ. Je suis bien conscient de ne pas avoir réussi à séparer alimentation et hydratation. Je suis toujours retourné à une équivalence. Une des questions initiales, qui était de savoir où va l'eau apportée par perfusion chez les patients en fin de vie qui ne s'alimentent plus, reste entière. Comment peut-elle retarder -un peu- leur mort ? Aucune explication explorée n'a donné de modèle présentable. La profonde unité du vivant exprimée non en terme de chimie (dicontinue, bottom-up...) mais en terme de fonctions (continues, top-down) permet d'intégrer facilement le discours philosophique à la compréhension des mécanismes du vivant, mais ne fournit pas de modèle utilisable expérimentalement. |
||||||||||||||||
|
|
||||||||||||||||||