|
|
|
|
|
|
||||
| |
Le programme nous demande de traiter 3 parties de façon plus ou moins séparée sans chercher à faire une étude intégrée de la fonction visuelle. Dont acte. La partie optique est traitée en physique et on peut s'appuyer sur elle. |
|
Sources: http://acces.ens-lyon.fr/acces/ressources/ neurosciences/vision/ comprendre/vision_scientifique
|
|||||
|
plan |
|
|
|
|||||
|
TP - Système nerveux (fait à la fin de la partie sur la reproduction) |
|
|||||||
|
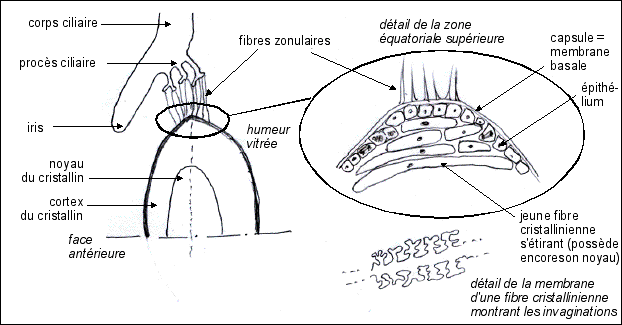
1 - Le cristallin : une lentille vivante |
|
|
||||||
|
TP- Anatomie de l'œil de thon et histologie du cristallin
|
|
Résumé |
|
Les cristallines représentent 90% des protéines hydrosolubles du cristallin - mais toutes les protéines, notamment les protéines membranaires ne sont pas hydrosolubles). |
||||
|
|
|
|||||||
| |
|
|
||||||
|
Transparence cristallinienne... Les tissus vivants sont souvent transparents et la plupart du temps translucides.
transparence =
capacité d'un milieu à laisser
passer une image nette (se différencie de la
translucidité = capacité
d'un milieu à laisser passer la
lumière, ce qui conduit à des
images floues ou des ombres, mais pas une
image nette ; s'oppose à l'opacité).
|
|
Les embryologistes utilisent pour leurs études des organismes modèles dont les embryons - au moins dans les premiers stades jusqu'à la fin de la morphogénèse - sont plus ou moins transparents, mais presque toujours translucides : Xenopus lævis Cænorabditis elegans poisson-zèbre (Danio rerio) |
|
L'opacité provient la plupart du temps d'accumulation de pigments dont la mélanine, chez les vertébrés est l'un des plus courants. Il ne faut pas oublier que l'on doit souvent colorer les coupes histologiques si l'on veut distinguer des structures au MO. Quelques mélanocytes de
l'épiderme de X. lævis (cellules de
l'épiderme synthétisant les grains de
mélanine - voir ancien
cours 1èreS) |
||||
|
... mais pas
d'ordre cristallin |
|
Mais les propriétés optiques du
cristallin vont bien au-delà de la simple
transparence : |
||||||
|
Vie ... L'absence de noyau empêche le renouvellement protéique par voie de synthèse, mais le métabolisme énergétique de la cellule nécessite une consommation de nutriments. |
|
Il faut distinguer deux types de
renouvellement: |
|
Le cristallin n'est ni innervé, ni vascularisé. Le glucose de l'humeur aqueuse est le principal nutriment des cellules du cristallin. Sauf dans les cellules épithéliales pourvues de mitochondries où la respiration est possible, l'oxydation du glucose est de type fermentaire (anaérobie, voir terminale). |
||||
| |
|
Les altérations du cristallin ont la plupart du temps une cause métabolique. Par exemple, chez les diabétiques l'excès de glucose circulant conduit à des réactions chimiques qui déstructurent les protéines cristalliniennes et provoquent l'opacification du cristallin (cataracte). |
|
|
||||
|
Déformation...
un super site: |
|
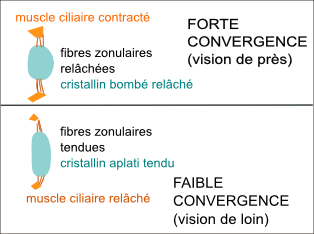
La cornée, du fait de
son bombement, constitue un appareil convergent, mais non
adaptable, alors que le cristallin par sa déformation
peut changer sa focale (ou sa vergence). Les propriétés d'élasticité du cristallin sont rapportées à sa composition en cristallines et filensines....! |
|
La déformation-accommodation est due
à la contraction du muscle ciliaire qui
est un petit muscle en anneau dont les fibres musculaires
sont elles-mêmes attachées au cristallin (au
niveau de la zone (ou zonule) de Zinn) par des prolongements
du corps cilaires apellées procès cilaires et
fibres zonulaires. |
||||
| |
|
Avec l'âge le cristallin perd en élasticité, ce qui conduit à des défauts d'accommodation apparaissant vers 45 ans (presbytie). |
|
|
||||
|
Remarque |
|
J'avais pensé, une
fois n'est pas coutume, faire plaisir aux tenants de
l'idéologie pédagogiste soixante-huitarde
fort répandue dans l'Education nationale et
construire mon cours sur une triple question (construite
à partir d'idées préconçues
que les élèves sont supposés
avoir...): Les réponses aux questions posées: - de nombreuses cellules vivantes sont transparentes, mais l'organisation cristallinienne est unique. - les cellules du noyau du cristallin (sans aucun organite) ont un métabolisme faible (fermentaire) et ne renouvellent plus aucun de leur systèmes structuraux ni enzymatiques. - le cristallin présente une déformation PASSIVE. En dernière
analyse, il est clair que ces questions sont finalement
mal posées et de peu d'intérêt. Le
caractère vivant n'est ni incompatible avec la
transparence, ni vraiment bien connu, voire défini
pour une cellule avec un métabolisme si faible.
Selon les fibres cristalliniennes les
propriétés sont variables. Ensuite, la
passivité du cristallin vis-à-vis de la
déformation ne rend pas compte des variations de
ses caractères physiques qui justement
déterminent son efficacité optique, qui est
l'objet de tant de recherches à visées
médicales. |
||||||
|
2 - Les pigments des photorécepteurs rétiniens et la vision des couleurs |
|
|
||||||
| |
|
Dans cette partie, le but du programme me semble tout autre : présenter une interprétation évolutive (plutôt phylogénétique) des pigments de la famille des opsines. C'est à mon avis prématuré en 1ère principalement à cause de l'étude du crossing-over qui ne se fait qu'en terminale. |
||||||
|
http://www.vetopsy.fr/sens/vision/transduction-visuelle-phototransduction.php Les pigments
photosensibles ont une partie protéique
variable : l'opsine et une partie commune : le chromophore.
Les pigments photosensibles sont des récepteurs (photoniques) membranaires couplés à des protéines G (au doux nom de transducine) qui activent un second messager (GMPcyclique).
|
|
Le rétinal (cis), lors de la réception d'un photon vibre (tout-trans) et provoque un changement de forme de l'opsine qui active la transducine qui transmet à d'autres molécules son excitation et notamment à une enzyme qui hydrolyse le GTP (en GMP) Le photorécepteur - au niveau du segment externe - est dépolarisé à l'obscurité et hyperpolarisé à la lumière. Les potentiels de récepteurs (potentiels locaux) apparaissent avec un seul photon (105 molécules de GMP sont formées à partir de la réception d'un seul photon !!!). Le neurotransmetteur reçu par la cellule ganglionnaire est le glutamate. |
|
Il y a 4 types d'opsine: 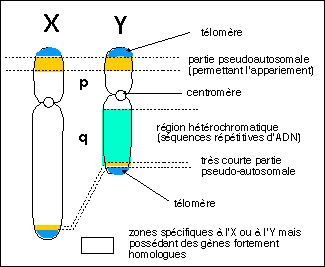 Grandes régions des chromosomes sexuels (gonosomes) schématisées |
|
La plupart des personnes sont trichromates
et possèdent les trois pigments. Les dichromates n'ont que deux pigments liés à la vision des couleurs fonctionnnels : le S (bleu) et soit le M (vert; protanopie) soit le L (rouge : deutéranopie). La plupart des Mammifères (commer le chien et le chat sont dichromates) (voir ici). |
||
|
Comparaison moléculaire des opsines : |
|
Le problème de fond de ces comparaisons moléculaires reste leur lien avec la biologie. La plupart des variants - apellés le plus souvent mutants - observés dans les différents taxons ne correspondent pas à des différences fonctionnelles. On IGNORE encore comment la séquence de l'opsine est reliée au spectre d'action (transduction lumière-potentiel cellulaire gradué) alors, relier à la fonction des variations concernant un seul acide aminé relève pour l'instant de l'espoir des chercheurs. Les arbres phylogénétiques construits sur ces données moléculaires peuvent parfois conforter des données évolutives anatomiques ou morphologiques mais il est peu prudent de les considérer comme un point de départ de la réflexion sur l'évolution. C'est un très mauvais point de départ pour l'évolution des Primates qui sera abordée en Terminale. |
|
On notera de plus que ces reconstitutions sont très dépendantes de certaines hypothèses concernant la méthode de comparaison de séquence... L'approche évolutive molécularo-génétique est fort limitée et très spéculative..La génétique permet de trouver des différences dans l'unité, mais elle rend peu compte de l'unité.Introduire l'évolution en 1ère S en se limitant à la séquence d'un pigment protéique est une mascarade du même type que l'importance exagérée donnée aux mutations dans la variation génétique et surtout dans la variation des espèces. L'anatomie comparée de l'œil, et particulièrement celui de l'œil des vertébrés permet une approche évolutive d'une toute autre richesse. |
|
|
||
|
Mécanismes chromosomiques rendant compte de l'organisation génomique des gènes des opsines |
|
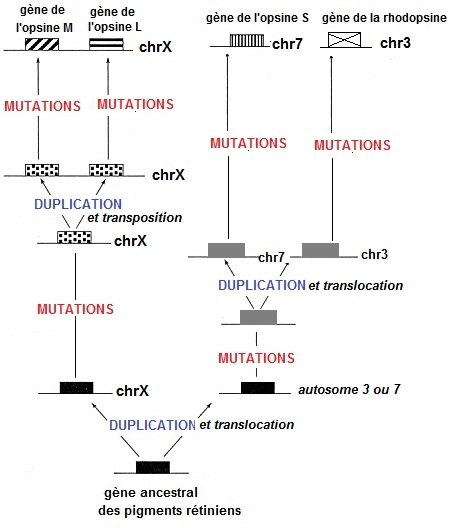 image externe |
|
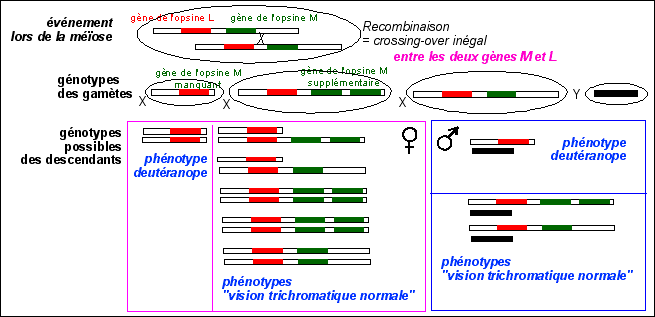
Il est par contre beaucoup plus pertinent de se limiter à des comparaison chromosomiques issues notamment des travaux de Jeremy Nathans dans les années 1980-1990. La proximité des gènes L et M sur le chromosome X (et leur similitude de séquence) peut faire penser à une origine par duplication de gène et peut nous aider à comprendre comment de nombreux défauts de la vision des couleurs prédominent chez les individus de sexe masculin (partie propre de l'X en un seul exemplaire alors que chez les femmes, cette partie de l'X est pseudoautosomale). |
||||
|
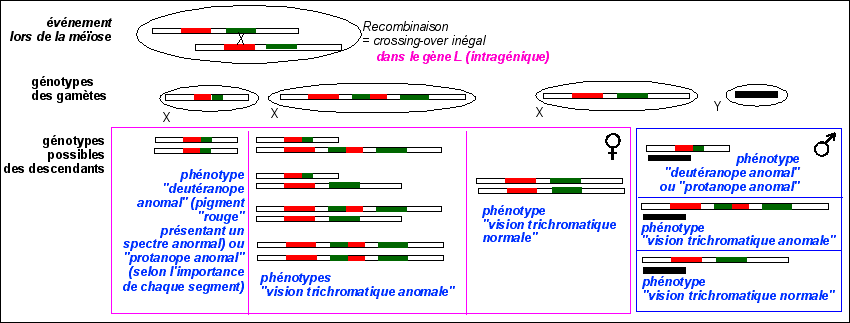
Interprétations chromosomique possibles de quelques anomalies de vision des couleurs
|
|
Un crossing-over inégal entre les deux gènes (M et L) pouvant donner lieu à des deutéranopes (un seul gène de type L) et des individus à vision des couleurs normale (une gène L et deux gènes M).
Le mécanisme évoqué ici (recombinaison de type crossing-ocer inégal) est un mécanisme possible de duplication du gène M, par lequel on a pu obtenir les gènes M multiples obervés (jusqu'à 5 exemplaires). |
||||||
|
Un crossing-over inégal à l'intérieur de l'un des deux gènes (M et L) pouvant donner lieu à des gènes non fonctionnels et donc à des trichromates deutéranomaux (L, M/L, M) et protanomaux (L/M, M).
|
|
|
||||||
|
3 - La rétine, un morceau de cerveau au fond de l'œil |
|
Le texte de cette partie emprunte largement
à la Conférence ENS Paris D'un abord nettement plus difficile mais
qui contient quelques éléments
intéressants: |
||||||
|
TP - Histologie de la rétine Les types cellulaires de la
rétine |
|
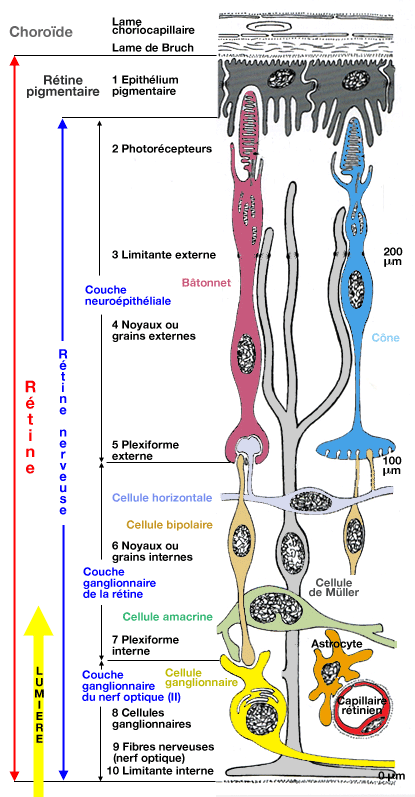
Dans la coupe de la rétine il est important de bien mettre en évidence les cellules épithéliales pigmentaires qui (i) empêchent un diffusion de la lumière d'un photorécepteur à l'autre (scattering) (ii) et qui phagocytent les disques du segment externe des cellules photoréceptrices qui sont produits en permanence (avec une durée de vie de l'ordre de 12 jours). On parle deux 2 rétines: la rétine pigmentaire et la rétine nerveuse. Penser aux cellules gliales (de Müller et astrocytes et microglie) ayant un rôle de soutien et de nutrition. |
|
La rétine, tant pigmentaire que nerveuse (voir TP), est vraiment un morceau de cerveau, même du point de vue embryonnaire du fait de son origine neurodermique. |
|
|
||
| |
|
|||||||
| |
Le travail expérimental sur la rétine (animale) extraite de l'œil et placée sur un support avec de nombreuses électrodes est FACILE. On projette des images directement sur cette rétine isolée et on récupére les signaux au niveau du nerf optique (fonctionne quelques heures). Au niveau des cellules photoréceptrices, des cellules horizontales et des cellules bipolaires les signaux électriques transmis sont des dépolarisations LOCALES (potentiels locaux) modulés en amplitude. Alors qu'au niveau des cellules ganglionnaires et des cellules amacrines, ce sont des potentiels d'action (PA) qui sont propagés (selon la loi du tout ou rien) et modulés en fréquence. |
Les cellules réceptrices sont très peu nombreuses. Par comparaison, il existe près de 1000 types cellulaires différents qui recoivent les molécules odorantes au niveau de l'épithélium nasal alors qu'il n'existe qu'un seul type de batonnets et que trois types de cônes. Si l'on élargi à l'ensemble des animaux on ne doit guère dépasser les 10 types cellulaires si l'on tient compte de la sensibilité aux u.v. par exemple. On a donc une stupéfiante UNITÉ et non pas une diversité. |
|
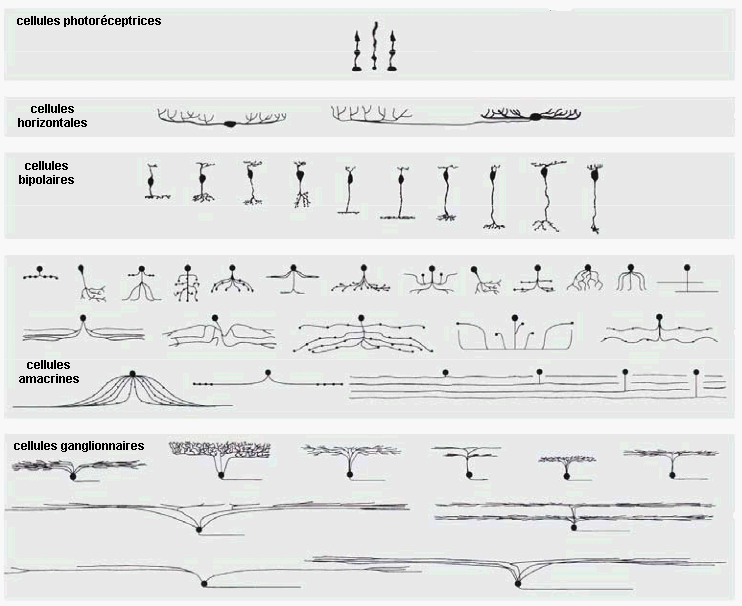
Par contre, l'organisation spatiale des
neurites de chaque type cellulaire est très
variée, ce qui est plutôt une indication de la
complexité du traitement rétinien des
informations visuelles : |
||||
|
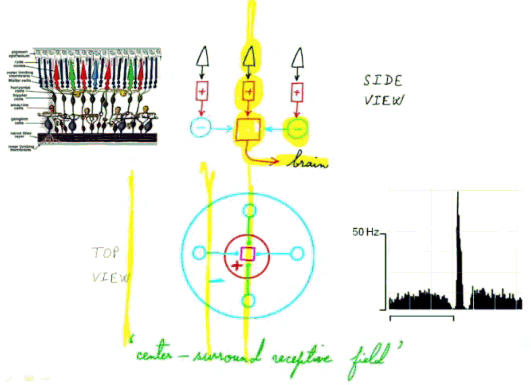
La rétine est organisée en champs récepteurs
|
|
Notre compréhension de l'organisation
de la rétine résulte grandement des
études faites sur des rétines isolées.
Les cellules photoréceptrices sont organisées
en champs récepteurs centre-pourtour
("center surround receptor
fields") de projection
globalement circulaire sur la rétine et comprenant de
quelques cellules à quelques dizaines de cellules (ce
qui fait des angles de vision qui vont d'une fraction de
degré à une dizaine de degrés selon les
animaux...) |
||||||
| |
Les champs récepteurs se recouvrent et l'on considère que chaque cellule ganglionnaire est le centre d'un champ récepteur. La taille des champs récepteurs. est plus étroite au centre du champ de vision et plus large sur les bords du fait de la disposition et du nombre de dendrites des cellules amacrines.
La tâche aveugle est le
lieu de départ du nerf optique et des vaisseaux
sanguins irriguant la rétine. Comme elle ne
possède pas de photorécepteurs elle correspond
donc à une zone sans information. |
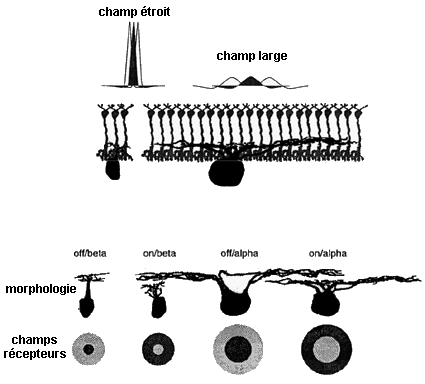 Diversité des champs récepteurs et relations topographiques entre cellules amacrines et bipolaires images extraites de Un regard sur la vision - Da Silveira Rava, ENS, 16 mars 2012 |
|
Test:  Clicquer sur l'image pour l'ouvrir seule dans une fenêtre |
||||
| |
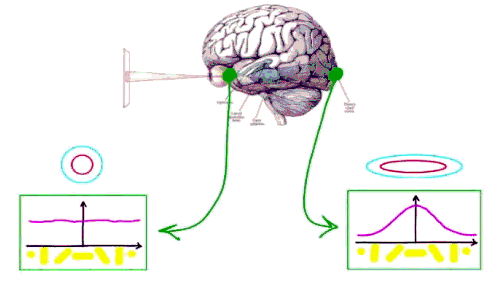
Dans les illusions d'optique il faut non seulement prendre en compte la sensibilité de la rétine à des différences de luminosité plutôt qu'à des intensité lumineuse mais il existe sans aucun doute des facteurs corticaux dans le traitement de l'information rétinienne. Dans le cortex les champs récepteurs
sont plus allongés et détectent une
orientation préférentielle
(tuning curve). |
|||||||
|
Quelques caractéristiques de la vision rétinienne |
|
|
|
|
||||
|
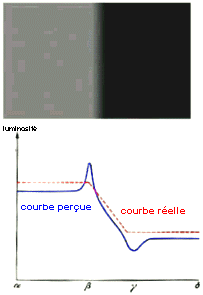
La rétine détecte des bords |
|
Les bandes de Mach sont des bandes d'intensité plus forte à la limitre des zones de changement de luminosité. Elles sont sans aucun doute dues à l'organisation en champ récepteurs : on détecte d'abord les bords !!! Dans l'image ci-contre on observe une bande
PLUS claire du côté clair et une bande PLUS
sombre du côté sombre alors que la
luminosité décroît de façon
régulière dans la partie centrale (courbe
rouge). La courbe de perception est en bleu. |
|
|
||||
|
La rétine détecte des différences |
|
Cette contrainte est sans aucun doute liée à l'organisation des champs récepteurs (on ne voit pas une intensité lumineuse absolue mais on détecte la différence d'excitation de plusieurs cellules. |
|
|
||||
|
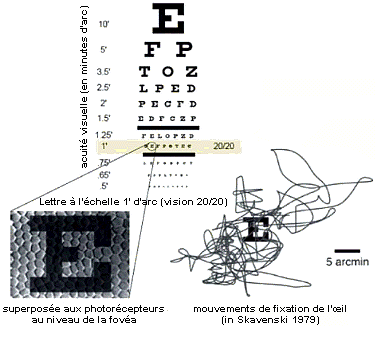
La rétine ne détecte qu'une image partielle mais qui nous semble complète
|
|
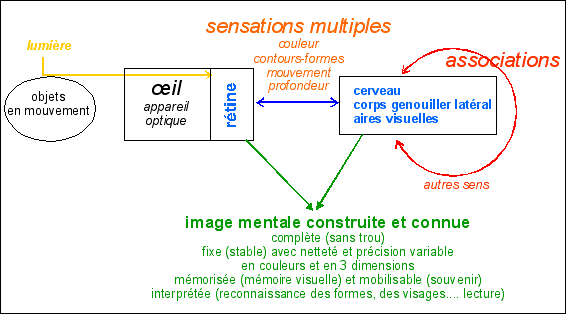
Si l'on peut dire qu'une image arrive à la rétine, cette image n'est pas transmise... seules les impulsions électriques (PA) sont transmises au cerveau. L'image rétinienne très partielle, car elle présente beaucoup de trous : vaisseaux sanguins, tâche aveugle (départ du nerf optique mais aussi arrivée des vaisseaux sanguins... bien visible sur l'image b ci-contre), fovéa sans bâtonnets (donc sans vision de nuit - centre noir de l'image d ci-contre)... Ce que le cerveau voit n'est donc pas une image mais bien une composition de multiples sensations (partielles) qui ont toute la rétine comme point de départ et qui son traitées dans les diverses zones nerveuses (rétine, corps genouillers latéraux (du thalamus), cortex visuel et autres parties du cortex...). De plus si la vision nécessite des traitements de sensations depuis la rétine vers le cerveau (sens montant = bottom-up) il existe sans consteste possible une modification de la sensation qui part du cerveau en direction de la rétine (sens descendant = top-down) et qui modifie la pérception visuelle à son origine. La rétine et le cerveau travaillent donc plus comme un peintre qui réaliserait plusieurs dizaines de représentations simultanées de la réalité. |
||||||
|
Les mouvements des objets et de l'œil sont indispensables à la vision.  Une fois que vous aurez vu - grâce au mouvement - ce qui se cache derrière ces points, vous le verrez à chaque fois que vous regarderez l'image, même sans mouvement. (extrait de la Conférence ENS Paris Un regard sur la vision - Da Silveira Rava, ENS, 16 mars 2012) Les petits mouvements involontaires de
l'œil qui servent à fixer l'image que l'on
regarde avec attention ("mouvements de fixation de
l'œil" = fixational eye movements) sont
intrinséquement liés à la
reconnaissance des objets puisqu'ils sont de plus grande
amplitude que les objets reconnus (voir ci-contre). Si l'on
stabilise l'image (œil absolument fixe) on devient
aveugle. Les illusions d'optique où l'on voit des mouvements dans des images fixes viennent de ce que le regard présente des mouvements propres d'assez grande amplitude qui trompent le système de reconnaissance d'une image fixe, la plupart du temps par l'absence de repère mémorisable dans l'image. Et puis bien sûr il y a le cinéma tout simplement qui est encore une fois une illusion de mouvement: les images fixes superposées créent l'illusion du mouvement (mouvement réel - caméra - images - projection - mouvement virtuel - rétine - cerveau). Certaines cellules rétiniennes sont spécialisées dans la direction du mouvement (droite gauche mais aussi approche ou éloignement et même isolement d'un mouvement particulier au sein d'autres mouvements). |
|
|
||||||
|
4 - La vision : du traitement rétinien d'images multiples en mouvement à la fonction cérébrale associative |
|
|
||||||
|
« Les informations provenant de la rétine sont à l'origine d'interactions entre de nombreuses régions du cerveau. Ces interactions aboutissent à la perception consciente de la scène visuelle, mais elles déclenchent en même temps des réflexes plus conventionnels comme l'ajustement du diamètre pupillaire, l'orientation des yeux vers les cibles intéressantes ou la régulation des comportemems homéostasiques liés au cycle nycthéméraL Les voies et les structures qui participent à ces fonctions sont nécessairement diverses. De toutes ces composantes du système visuel, la voie visuelle primaire qui va de la rétine au corps genouillé latéral du thalamus et, au-delà, jusqu'au cortex visuel primaire, est certainemem la plus importante et celle qui a fait l'objet des recherches les plus approfondies. Au sein de cetre voie, différentes catégories de neurones codent les aspects de l'information visuelle qui constituent ce que nous percevons finalement: luminance, différences spectrales, oriemation, mouvement. Le traitement parallèle de ces catégories d'information se poursuit dans des filières corticales qui s'étendent au-delà du cortex primaire et qui alimentent tout un ensemble d'aires visuelles réparties dans les lobes occipitaux, pariétaux et temporaux. Les aires visuelles du lobe temporal sont principalement impliquées dans la reconnaissance des objets, celles du lobe pariétal dans ce qui concerne le mouvement. Une vision normale dépend de l'intégration des informations de toutes ces aires. Les processus qui sous-tendent la perception visuelle ne sont pas élucidés et demeurent l'un des défis majeurs des neurosciences modernes. » Neurosciences, Purves et col., 4ème ed., 2011, De Boeck, p 289 |
|
|
||||||
|
L'idée principale serait que le cerveau, loin de restituer une image en deux dimensions qui s'imprimerait localement au niveau d'une surface plus ou moins corticale, traiterait de façon complexe et simultanée, les très nombreuses sensations visuelles de l'objet perçu, restituées par les différents types de groupements de cellules rétiniennes. Un
bon cours notamment sur la partie physio-psychologie : |
|
|
||||||
|
Une chaîne neuronale nutritive.... des associations non pas uniquement informatives mais beaucoup plus étroites |
|
La technique de marqage par des acides aminés radioactifs (aa*) a donné des résultats inatendus (Grafstein, 1971, 1975, Cowan et al. 1972) depuis que l'on s'est rendu compte que les aa* absorbés au niveau d'un corps cellulaire par exemple ganglionnaire rétinien étaient non seulement incorporés aux protéines transportées en direction des terminaisons synaptiques mais aussi transportés (en faible quantité) de la zone pré à la zone postsynaptique. On a ainsi pu marquer radioactivement (sur des mois d'expérience) des chaînes neuronales complètes depuis la rétine jusqu'au cortex visuel chez le chat et le singe. |
||||||
|
Une plasticité fonctionnelle intracellulaire |
|
Pour ce qui concerne la plasticité synaptique il faut aussi prendre en compte les nouvelles compréhensions de la membrane qui reviennent de façon extrémement fine (on suit les molécules une à une) à la notion de mosaïque fluide. Avant la biologie moléculaire se faisait sur du matériel mort, isolé, brotyé, solubilisé.... maintenant on suit les molécules unes à unes in vivo. Les zones post-synaptiques sont structurées par des protéines intracellulaires sous-membranaires et les récepteurs se déplacent (par diffusion ...) dans la membrane en établissant des connexions avec les protéines sous-membranaires ou d'autres interactions moléculaires qui déterminent des champs de récepteurs DYNAMIQUES. (cf conf Antoine Triller, ENS, académicien, "Une révolution en biophysique", 2009). On a accès aux mouvements des molécules in vivo et on a aussi accès aux constantes chimiques (concentrations, potentiels chimiques, temps de résidence, coefficients de diffusion...). |
||||||
|
Cerveau et vision : aires cérébrales et plasticité Plusieurs aires corticales participent à la vision. L'imagerie fonctionnelle du cerveau permet d'observer leur activation lorsque l'on observe des formes, des mouvements. La reconnaissance des formes nécessite une collaboration entre les fonctions visuelles et la mémoire. Des substances comme le LSD perturbent le fonctionnement des aires cérébrales associées à la vision et provoquent des hallucinations qui peuvent dériver vers des perturbations cérébrales graves et définitives. La mise en place du phénotype fonctionnel du système cérébral impliqué dans la vision repose sur des structures cérébrales innées, issues de l'évolution et sur la plasticité cérébrale au cours de l'histoire personnelle. De même la mémoire nécessaire par exemple à la reconnaissance d'un visage ou d'un mot repose sur la plasticité du cerveau. L'apprentissage repose sur la plasticité cérébrale. Il nécessite la sollicitation répétée des mêmes circuits neuroniques. Objectifs et mots clés. On cherche à montrer comment la réalisation d'une fonction cognitive complexe repose sur l'activité de plusieurs zones cérébrales de façon coordonnée. La maturation des systèmes cérébraux liés à la vision permet, à partir d'un nombre limité d'exemples, de présenter la notion de plasticité cérébrale et son lien avec l'apprentissage. [Limites. Il ne s'agit pas de présenter une étude exhaustive des aires cérébrales intervenant dans la vision, ni des mécanismes précis de la mémoire ou du langage. La notion de plasticité cérébrale est abordée à partir d'un nombre limité d'exemples, sans souci d'exposé exhaustif de ses mécanismes] Exploiter des données notamment expérimentales pour comprendre qu'une image naît des interactions entre différentes aires du cortex cérébral. Recenser, extraire et organiser des informations pour comprendre le phénomène de plasticité cérébrale et son importance dans l'établissement de différentes fonctions cognitives. Interpréter des expériences sur la maturation du cortex visuel chez l'animal. Interpréter des observations médicales et/ou des imageries cérébrales chez l'Homme. |
||||||||