II. Mécanismes de
l'immunité (1ère
partie)
Nouveau
cours 2008 :
généralités
- SIDA
-VIH
Tout ce chapitre correspond à une refonte des
éléments du programme dans un souci de cohérence
avec la conception de la vie présentée dans le cours de
seconde.
1 . A la recherche de l'identité de l'homme ;
2 . L'homme, milieu de vie ;
3 . L'homme blessé ; 4 . L'homme
malade; 5 . L'homme sain
TP: le système immunitaire: des
cellules et des organes -
TD: retour sur les cellules et les
molécules du système immunitaire
retour sommaire cours Terminale
S
Remarque: les numéros de pages font
référence au livre de TS de R. Tavernier et C. Lizeaux,
Bordas, enseignement obligatoire
Le document vidéo "La peau" de l'APBG permet de visualiser de
magnifiques cultures de kératinocytes qui illustrent
très bien la mobilité des cellules...
l'équivalent pour les cellules immunitaires du film d'Eric
Périlleux chez Nathan: "Le système immunitaire et la
communication entre cellules" qui, malgré ses longeurs, peut
être un excellent support à la présentation du
système immunitaire comme système de communication
(uniquement pour ses images de
microcinématographie).
Etymologiquement (Petit Robert, 1984) il semblerait que le terme
d'immunité vienne du latin immunitas:
exempté de charge (munus), sens datant de 1276: plus
récemment (1866) l'immunité recouvre les
mécanismes de défense d'un organisme vis-à-vis
d'un organisme pathogène (du grec pathos=malheur ou
souffrance). D'une façon plus générale,
l'immunité biologique semble avoir pris maintenant un sens
positif et correspondre actuellement à l'ensemble des
mécanismes développés par un organisme pour se
maintenir en bonne santé (immun dans le sens de sain). La
définition de l'encyclopédie Hachette multimédia
par exemple (1996) est caractéristique : l'immunité est
la "propriété que possède un organisme vivant
de développer des moyens spécifiques de défense
(naturels ou acquis) contre un agent pathogène
extérieur (infectieux, toxique, tumoral) ou contre un corps
étranger (greffe, cellule d'un autre individu)". Ou encore
celle d'"Immunologie" de Revillard (De Boeck Université,
1995): "L'immunité est l'ensemble des mécanismes
biologiques permettant à un organisme pluricellulaire de
maintenir la cohérence des cellules et tissus qui le
constituent, et d'assurer son intégrité en
éliminant les substances étrangères et les
agents infectieux auxquels il est exposé". Ce genre de
définition à rallonge prouve que le domaine de
l'immunité est mal défini. Dans un contexte encore plus
général, l'immunité doit être
traitée comme faisant partie des relations entre
espèces (relations inter spécifiques) qui,
elle-même, font partie du travail de relation des êtres
vivants (voir fiche "diversité du
vivant").
On a donc d'une part l'aspect "défense-maintien de la
cohérence-survie" vis-à-vis d'une agression et d'autre
part, maintien de l'"intégrité". Ces deux questions ne
sont pas forcément liées. En ce qui concerne
l'intégrité, il faut avant tout définir
l'identité de l'organisme. Dans ce chapitre nous ne traiterons
que l'immunité humaine mais il est évident que la
plupart des concepts proposés peuvent être
élargis à d'autres organismes pluricellulaires (y
compris les végétaux (plantae) pour lesquels les
connaissances sont encore peu nombreuses.. du moins pour ce que j'en
sais).
Une bonne part des concepts immunitaires vient de
résultats d'expériences réalisées chez
des animaux de laboratoire, essentiellement des Mammifères
mais on connaît un système immunitaire chez tous les
vertébrés. Dans quelle mesure les connaissances
acquises chez ces animaux sont extrapolables chez l'homme: c'est
encore une fois les résultats des pratiques médicales
qui nous l'indiquent mais il faut être prudent et penser que
l'on travaille souvent avec un "modèle animal" et non avec des
résultats obtenus chez l'homme.
Pour une petite entrée sur la notion de maladie et de
santé voir une page à l'attention
des élèves de terminale curieux.
1. A la recherche de
l'identité de l'homme
Comme nous l'avons présenté dans le cours de seconde
les trois niveaux du vivant sont inséparables. On distingue
alors une identité génétique qui se
traduit par une identité moléculaire, ce que le
programme résume par le terme "marqueurs du soi". Il existe
ensuite une identité cellulaire (qui détermine
aussi une identité tissulaire et même organique qui
dépasse le simple niveau moléculaire); c'est le domaine
d'étude des greffes par exemple ou des cancers. Enfin, on
distingue le niveau de l'organisme entier pour lequel on peut parler
d'identité biologique. C'est à se niveau que
l'on peut étudier par exemple l'intégration de
l'individu au sein de l'écosystème, ses moyens de
défense nécessaires à sa survie... etc. Par
souci de "coller" au programme nous ne pourrons développer de
façon harmonieuse chaque partie mais on s'efforcera tout de
même de balayer la question le plus largement possible.
- L'identité biologique
Tout individu peut être reconnu par des traits
qui lui sont spécifiques, des traits matériels
(forme du nez, empreintes digitales, forme des oreilles...) ou
comportementales (voix, gestes...). En l'état actuel de nos
connaissances ces traits personnels ne sont pas réductibles
à un déterminisme ni génétique, ni
cellulaire. L'inégalité des humains devant la
maladie est un élément incontournable et il est
simpliste et même dangereux de vouloir expliquer toutes ces
individualités par des différences
génétiques. Chaque individu participe à la
construction de sa propre personnalité sans qu'elle soit
déterminée de façon absolue. La
résistance de chacun à différentes maladies
dépend d'une part de son histoire, des
différents contacts qu'il a pu avoir avec différents
antigènes (ou éléments déclenchant une
réaction immunitaire), et d'autre part du terrain
biologique, c'est-à-dire d'un ensemble d'interactions
complexes qui font qu'il présente plus ou moins telle ou
telle aptitude à la défense contre tel ou tel
antigène.
- L'identité cellulaire, tissulaire et
organique
L'identité au niveau des cellules et des tissus
est essentiellement étudiée à partir de
greffes et surtout de greffes sanguines (en effet le sang est un
tissu... liquide). Cette identité est un des aspects de la
différenciation, thème trop vaste pour être
abordé ici.
|
GREFFE
|
Une greffe est le prélèvement
d'un tissu ou d'un organe chez un donneur et sa
transplantation chez un receveur sans établir de
chirurgicalement connections (anastomoses) vasculaires
(par exemple: la peau, la cornée, la moelle
osseuse...). On parle d'autogreffe lorsque le
donneur et le receveur sont le même individu, de
greffe syngénique lorsque ce sont de vrais
jumeaux (monozygotes), d'allogreffe lorsqu'ils
sont de la même espèce et de
xénogreffe lorsqu'ils sont d'espèces
différentes.
|
|
TRANSPLANTATION
|
Une transplantation est le
prélèvement d'un tissu ou d'un organe chez
un donneur et sa transplantation chez un receveur avec
rétablissement chirurgical de la continuité
vasculaire (rein, coeur, foie, pancréas, poumon,
coeur-poumon...).
|
Voici quelques expériences qui pourront aider à
présenter les différents niveaux d'étude.
|
Expériences
|
pistes
d'interprétations
|
|
1
|
Une greffe de peau (épiderme et derme)
réalisée entre deux souris issues de
parents différents est immanquablement suivie d'un
rejet plus ou moins rapide (le rejet se
caractérise par une mort du greffon du à
une absence de vascularisation, et à
l'infiltration du greffon par des cellules immunitaires
(lymphocytes, macrophages) qui provoquent la mort des
cellules du greffon).
|
Il y a reconnaissance et destruction d'une peau
étrangère par le système immunitaire
du receveur
|
|
2
|
Une greffe d'épiderme seul sans cellules de
Langerhans (uniquement les kératinocytes
cultivés in vitro) réalisée
entre deux individus issus de parents différents
est , en principe, non rejetée.
|
Les kératinocytes cultivés in vitro
ne sont pas reconnus comme étrangers: soit ils
perdent leur spécificité lors de leur
culture, soit ils ne possédent pas de marqueurs de
l'identité reconnue lors de greffes; ce qui peut
s'exprimer ainsi: en absence d'élément du
système immunitaire du donneur il n'y a pas de
reconnaissance d'un tissu par le système
immunitaire du receveur.
|
|
3
|
Chez des souris mutantes qualifiées de nude
(nu//nu) et dépourvues de thymus fonctionnel
(entre autres anomalies...) ou chez qui on supprime le
thymus dès la naissance et que l'on irradie enfin
fortement (l'irradiation provoque la destruction des
cellules souches immunitaires de la moelle osseuse), une
allogreffe de peau est toujours acceptée.
|
La reconnaissance de peau étrangère
et son acceptation ou son rejet par la souris est sous le
contrôle simultané et indissociable de deux
organes essentiels : le thymus et la moelle
osseuse.
|
|
4
|
Si l'on déplace certains tissus chez un
même individu (comme par exemple du tissu
musculaire) et qu'on les greffe au sein d'une autre
structure (par exemple entre les couches de
l'épiderme), le greffon
dégénère. Les tissus greffés
sont rapidement détruits par des cellules
immunitaires de type lymphocytes et phagocytes (du grec
phagein = manger).
|
Il existe des mécanismes de reconnaissance
de lignées cellulaires différenciées
au sein de l'organisme. On ne peut pas ajouter des
cellules différenciées d'une autre
lignée à une population cellulaire
homogène.
|
L'identité cellulaire passe donc par la reconnaissance, par
le système immunitaire, de caractéristiques
cellulaires qui peuvent aussi bien être cytoplasmiques que
génétiques. L'identité cellulaire qui passe
par la mémoire cytoplasmique n'est pas qu'une simple
suggestion sans aucun fondement expérimental. Elle
résulte de la profonde unité du vivant que nous
avons abordé au sujet des mécanismes du vivant
(la vie au travail). Les
études expérimentales embryologiques
réalisées par le Professeur Rosine Chandebois
permettent d'en saisir les principes qui sortent cependant du
cadre de ce cours. Je ne peux que vous suggérer de vous
reporter à ses ouvrages (voir bibliographie,
notamment "Le gène et la forme").
Remarques :
- On lit parfois que les grossesses
sont des cas d'allogreffes qui sont tolérées
pendant un certain temps.
Une suggestion qui change tout:
En terme d'identité cellulaire le fœtus n'est
PAS une allogreffe. Étant donné la faible
participation du cytoplasme du spermatozoïde à la
fécondation, on peut affirmer que l'identité
cytoplasmique de la cellule œuf est avant tout celle de
l'ovocyte et donc celle d'une cellule maternelle.
L'identité cellulaire du fœtus est celle de sa
mère. Il y a alors autogreffe et non
allogreffe.
Remarque:
Mes élèves me font remarquer à juste titre
qu'ils sont choqués par le terme de greffe. La
participation essentielle de la cellule sexuelle maternelle
(ovocyte II) à la conception et la profondeur de ce
travail de reproduction intimement lié à la
dimension maternelle de la nature féminine, ne sont pas
pris en compte dans ce mot. La gestation en terme immunitaire
devrait à mon sens être pris d'une part comme une
éducation du système immunitaire de l'enfant par
celui de sa mère et d'autre part en terme de protection
: la grossesse me semble être avant tout un
développement protégé d'un embryon au sein
de l'organisme maternel. Le terme de parasite que l'on applique
parfois au foetus n'est aussi qu'un pâle reflet de
l'originalité des relations entre la mère et
l'enfant.
Interprétation : la "cohabitation" des
systèmes immunitaires maternel et foetal,
qualifiée de façon excessive de tolérance,
est largement inexpliquée actuellement si l'on ne
raisonne qu'en terme d'identité génétique.
Dans ce cas en effet le fœtus possède à la
fois l'identité paternelle et maternelle. Mais il est
probable que l'on fasse fausse route. L'identité
immunologique cellulaire étant celle de la mère,
il n'y a plus aucun problème sauf justement pour
l'identité génétique (voir ci-dessous).
Des hématies fœtales passent dans le sang de la
mère en fin de grossesse et lors de l'accouchement.
Elles induisent la production d'anticorps (substances
défensives sécrétées par les
plasmocytes comme nous le verrons plus tard) dirigés
contre les antigènes (marqueurs membranaires) des
globules rouges du fœtus. C'est ce qui intervient
notamment lorsque la mère est de groupe sanguin
rhésus négatif et que son enfant est de groupe
rhésus positif.
- Le développement des techniques
moléculaires fait parfois oublier les
caractéristiques mécaniques d'une cellule. Par
exemple si l'on étudie le vieillissement des
hématies. Il est certes très intéressant
de rechercher des marqueurs moléculaires membranaires du
vieillissement qui seraient reconnus par les macrophages, mais
il est tout aussi intéressant de s'intéresser aux
propriétés mécaniques d'une hématie
vieillissante : perte d'élasticité tout
simplement amenant une moindre déformation et
peut-être le blocage au niveau d'un capillaire
étroit (comme certains capillaires de la rate qui jouent
le rôle de filtres)... De la même façon, le
gonflement d'une cellule activée peut expliquer un
changement de surface qui puisse être reconnu par un
autre type cellulaire ou tout simplement encore son blocage au
niveau d'un capillaire lymphatique étroit la mettant en
contact avec d'autres cellules.
- L'identité génétique,
support de l'identité moléculaire
Les immunologistes, toujours dans le cadre des
interprétations de greffes, principalement sanguines, se
sont dotés des outils du génie
génétique (voir cours de
spécialité). Le but étant de
TROUVER des MARQUEURS MOLÉCULAIRES DE
L'IDENTITÉ DE L'ORGANISME, c'est-à-dire des
molécules suffisamment variables d'un individu à
l'autre pour être originales chez chacun, mais pourtant
constantes chez toutes les cellules d'un même
organisme.
Parmi les nombreuses substances chimiques qui différent
entre le donneur et le receveur d'une greffe, on a très
tôt (années 1950) reconnu un groupe
d'antigènes tissulaires codés par des gènes
polyalléliques définissant les systèmes
d'histocompatibilité. Par exemple le système des
groupes sanguins ABo (de Landsteiner) correspond à des
molécules tissulaires (ici le tissu est un peu particulier,
c'est un liquide : le sang) très fortement
antigéniques.
Les groupes sanguins de
Landsteiner - le système ABo
(Karl Landsteiner (1868-1943) découvrit
les groupes sanguins ABo en 1900 et le système
rhésus en 1940)
|
|
Cellules:
uniquement les globules rouges (hématies)
humains (?)
|
|
Molécules antigéniques: (cf
Bordas TS, p 88 n°5 )
Glycolipides ou glycoprotéines
insérés dans la membrane des
hématies: le marqueur A diffère du marqueur
B par un N-acétyl-glucosamine à la place
d'un galactose; le marqueur o ne possédant aucun
de ces deux sucres à l'extrémité de
sa chaîne glucidique. Ces éléments de
la chaîne glucidique sont accrochés par des
enzymes aux molécules porteuses au sein de
l'appareil de Golgi et du réticulum endoplasmique
lisse lors de la maturation des composants
membranaires.
|
|
Gènes:
les groupes sanguins ABo semblent
déterminés par un gène lié
au chromosome 9 et présentant trois
allèles. Les marqueurs A et B peuvent
être exprimés simultanément chez les
individus de groupe AB, on dit que ces caractères
sont codominants. Le caractère o est
récessif vis à vis des deux autres
marqueurs : il peut être présent à la
fois avec les marqueurs A ou B mais n'apparaît que
lorsqu'il est seul (homozygote o//o).
|
|
Réaction antigénique:
des sangs possédant les marqueurs A et B seuls ne
peuvent se mélanger sans que l'on observe une
réaction d'agglutination (les
hématies se collent les unes aux autres et forment
de petits amas). Seuls sont miscibles (compatibles) des
sangs de même groupe (transfusion isogroupe) ou de
o vers n'importe quel receveur (donneur universel). Le
groupe o ne peut recevoir que du groupe o sous peine
d'agglutination du sang transfusé (accident
transfusionnel extrêmement fréquent avant la
découverte des groupes sanguins et du respect des
lois de transfusion précisées ci
après). Le groupe AB est le receveur universel car
il n'agglutine pas les hématies du donneur quelque
soit son groupe. Les substances chimiques du sérum
responsables de l'agglutination sont les anticorps
ou agglutinines. Elles sont
sécrétées par l'organisme en
réponse aux marqueurs sanguins non présents
chez le receveur : un individu de groupe A pouvant
sécréter des anticorps anti-B et un
individu de groupe B des anticorps anti-A. Les individus
de groupe AB ne sécrètent ni anti-A ni
anti-B et les individus de groupe o peuvent
sécréter les deux types d'agglutinines. On
a pu ainsi réaliser par le passé des
transfusions sanguines entre groupes différents en
notant que les accidents transfusionnels (agglutinations)
dépendaient surtout des marqueurs du donneur et
des agglutinines du receveur. Effectivement, on peut
imaginer que le sérum du donneur, et donc ses
agglutinines, sont diluées dans celui du receveur
et donc incapables d'agglutiner les hématies de ce
même receveur. Actuellement , on ne fait plus que
des transfusions isogroupes. Les anticorps anti A et anti
B sont produits par les lymphocytes B comme nous le
verrons plus tard.
Remarque :
Normalement les anticorps ne sont produits qu'en
présence d'un antigène. Les groupes
sanguins semblent constituer une exception. On pense que
les anticorps seraient sécrétés en
réponse à des antigènes des
micro-organismes de la flore intestinale... L'absence
d'anticorps dirigés contre les antigènes de
ses propres hématies n'est pas non plus un fait
clairement expliqué. Les théories
présentant les différents clones de
lymphocytes supposés présents dans
l'organisme ont été retirées avec
raison du programme.
|
Le système d'histocompatibilité le plus connu est le
complexe majeur d'histocompatibilité (CMH), les autres
étant qualifiés de
mineurs.
Le CMH (complexe majeur
d'histocompatibilité)
Une difficulté de vocabulaire bien
lourde à traîner : le terme de CMH
désigne un groupe de gènes. Les
antigènes (molécules marqueurs de surface
des cellules) sont donc des produits du CMH. MAIS
CE NE SONT PAS LES SEULS. Il est donc interdit de parler
des molécules du CMH pour désigner
les seuls produits antigéniques de surface; par
contre il est très fréquent de parler des
molécules HLA, ce qui est pourtant tout autant un
abus de langage.
|
|
Découverte:
C'est Jean Dausset qui, en 1958, fait la première
description de ces antigènes à la surface
des globules blancs sanguins (leucocytes) à partir
de l'analyse de réactions d'agglutination obtenues
avec des sérums de sujets immunisés
à l'occasion de transfusions sanguines. Le
système de gènes codant pour ces
antigènes a été depuis lors
étudié en détail et nommé
HLA (Human Leucocyte Antigens). Découvert
aussi chez d'autres espèces de Mammifères,
notamment chez la souris où il est appelé
H2, on le qualifie de façon générale
de CMH ou complexe majeur
d'histocompatibilité.
|
|
Gènes et produits:
Les gènes du CMH chez l'homme (système HLA)
forment un groupe multigénique,
multiallélique et d'expression
codominante. Ils sont localisés sur le bras
court du chromosome 6. Ils codent pour des marqueurs de
surface de nombreuses cellules mais aussi pour des
enzymes, pour certains éléments du
complément (ensemble de protéines du
sérum intervenant dans la réaction
immunitaire comme nous le verrons plus loin), pour des
molécules intervenant dans la communication entre
cellules immunitaires ou encore pour des enzymes
intervenant dans le métabolisme de la cellule.
Leurs produits ne sont donc pas tous des molécules
intervenant dans la seule immunité... Le nombre de
gènes connus est voisin de 30 (d'où le
terme de multigènique). Pour chaque gène
connu la liste des allèles détectés
par des méthodes utilisant les sérums ne
cesse de croître. Elle peut aller d'une dizaine
à plusieurs dizaines d'allèles (famille
multiallélique). Cette variabilité est
qualifiée de polymorphisme : on recense plus de
1010 combinaisons allélique possibles
chez un individu. Il existe d'autre part des
mécanismes originaux de conversion génique
par échange de séquence d'ADN entre
gènes qui ont été mises en
évidence lors de la maturation de certaines
cellules...Bref tout cela est fort complexe.
|
|
Les molécules HLA,
antigènes membranaires
|
|
Antigènes :
Ce sont des glycoprotéines formées
de deux chaînes . Leur structure quaternaire
(disposition dans l'espace) est connue pour la plupart
d'entre elles.Les molécules HLA que l'on trouve
à la surface de la majorité des cellules
comportent une chaîne invariable (ß2
microglobuline) insérée dans la membrane et
liée de façon non covalente à une
chaîne lourde (a)
polymorphique située à l'extérieur
de la cellule et qui présente une cavité
dans laquelle peuvent venir s'insérer de petits
peptides d'environ 9 acides aminés. Les
molécules HLA sont formées de deux
chaînes polymorphiques (a,
b) comportant chacune deux
domaines extra-membranaires présentant aussi une
cavité extérieure à la cellule et
pouvant contenir de petits peptides. Une chaîne
légère invariable (li) est associée
aux deux chaînes précédentes sur la
face interne de la membrane.
|
|
Cellules :
Un premier groupe de molécules HLA (classe I) est
présent à la surface de la plupart des
cellules nucléées de l'organisme
(104 à 5x105
molécules par cellules mais pratiquement
absentes des hématies et des cellules nerveuses,
des cellules des glandes salivaires ou encore des
cellules embryonnaires au début du
développement...). Un autre groupe (classe II) est
formé de molécules HLA dont la
présence est strictement limitée
à certaines cellules de l'organisme intervenant
dans l'immunité (105
molécules par cellule par exemple pour les
lymphocytes , les phagocytes, les cellules de
l'épithélium (tissu de revêtement)
intestinal ou encore de l'épithélium
respiratoire. Cette répartition prouve clairement
que les molécules HLA ne peuvent pas être
considérées comme des déterminants
exclusifs du soi, compris comme identité
biologique (ni de l'identité moléculaire
d'ailleurs).
|
|
Gènes :
La chaîne invariante légère (ß2
microglobuline) des molécules HLA du premier
groupe est codée par un gène, qui ne fait
pas partie du CMH, et présent sur le chromosome
15. La chaîne lourde (a)
est codée habituellement par 3 gènes du CMH
et donc situés sur le chromosome 6. Les
chaînes polymorphes des molécules HLA du
second groupe sont codées par des gènes du
CMH, entre 23 et 25, regroupés en trois
régions (DR, DP et DQ). La chaîne invariable
li est codée par un gène localisé
sur le chromosome 5.
Il existe un déséquilibre de liaison
spécifique à certaines populations. En
effet, on observe des différences entre groupes
ethniques, certains allèles se retrouvent plus
fortement associés dans certaines populations que
ne le voudrait une distribution aléatoire....
C'est encore une preuve de la manipulation des
gènes par la cellule et du fait que l'on travaille
toujours sur une interprétation chromosomique de
la liaison de tel ou tel caractère...
|
|
Rôles :
Les molécules HLA du premier groupe (classe I)
sont supposées être responsables de la
présentation des peptides (9 aa) issus de la
dégradation permanente par chaque cellule de
ses propres protéines (protéines
endogènes). La cellule présenterait
ainsi en surface en permanence des fragments des
protéines qu'elle synthétise. Ces
protéines seraient bien sûr modifiées
dans le cas d'une infection virale, la cellule
synthétisant alors les protéines
virales.
Les molécules HLA du second groupe (classe II)
sont supposées présenter des peptides (9
aa) qui sont issus du milieu extérieur, ont
été absorbés par la cellule
(endocytose) puis dégradés.
Il semblerait qu'il existe un seuil à partir
duquel la présentation des peptides provoque une
réaction immunitaire. Il serait d'environ de 200
molécules HLA associées au même
peptide. La plupart des peptides présentés
par les molécules HLA sont des fragments de ces
mêmes molécules HLA. De nombreuses
séquences de peptides sont identiques (homologues)
à celles de peptides de micro-organismes. Enfin,
si le nombre théorique de peptides possible est de
5.1011 (9 aa parmi 20 aa) , il semble que le
nombre réél est beaucoup plus restreint du
fait de leur aptitude ou non à établir des
liaisons avec les molécules HLA (essentiellement
par 2 aa de la cavité des molécules
HLA).
|
Déterminer les différents allèles pour les
systèmes d'histocompatibilté s'appelle faire un
groupage ou typage ABO ou HLA par exemple.
En conclusion: on est encore assez
loin d'avoir une idée très précise de
l'identité moléculaire d'une cellule, d'un organe et
encore moins d'un organisme. Les produits du CMH se sont
avérés être très complexes et si
leur rôle dans le déclenchement de la
réponse immunitaire spécifique devient de plus en
plus clair (voir plus loin), leur rôle comme marqueur
du soi moléculaire devient par contre de plus en plus
flou.
|
En résumé: (à
illustrer à l'aide du schéma Bordas p 101
mais, attention, les 3 cas présentés sont
ceux des "molécules du CMHI)")
* des glycoprotéines membranaires dites
"molécules du CMHI" (car
associées pour une partie de leurs chaînes
protéiques aux gènes du CMH de classe I
portés par le chromosome 6) présentent
en permanence à la surface des membranes de
presque toutes les cellules de l'organisme des fragments
(9 aa) des peptides que la cellule fabrique (peptides
endogènes). (Les cellules qui ne portent presque
pas de ces molécules sont les hématies, les
cellules nerveuses, les cellules embryonnaires...). On
parle abusivement dans les manuels scolaires de
"présentation du soi ou du soi
modifié" (si l'on pense aux
molécules du CMH modifiés par les peptides)
pour les cellules synthétisant des
protéines habituelles et de "présentation
du non-soi" pour des cellules synthétisant des
protéines anormales. Il serait plus juste de
parler de présentation de l'activité de
synthèse des protéines de la cellule. On le
comprend comme un moyen d'exposer pour chaque cellule
un reflet de son activité de synthèse
protéique, afin que d'autres cellules, les
cellules immunitaires, puissent détecter la
présence de peptides anormaux. La surveillance
immunitaire étant dirigée vers presque
toutes les cellules nucléées.
* des glycoprotéines membranaires dites
"molécules du CMHII" (car
associées pour une partie de leurs chaînes
protéiques aux gènes du CMH de classe II
portés par le chromosome 6) présentent
en permanence à la surface des membranes de
presque toutes les cellules immunitaires
(lymphocytes, phagocytes, cellules du système
lymphoïde associé aux muqueuses) des
fragments (9 aa) des peptides captés dans le
milieu intérieur de l'organisme (peptides
exogènes), la cellule les internalisant
(endocytose) et les fragmentant. On ne dissocie pas ce
rôle dans les manuels scolaires de la
"présentation du non-soi", ce qui est dommage.
Ceci est une véritable activité de
surveillance ou de veille des cellules immunitaires
dirigée vers les protéines circulant dans
le milieu intérieur.
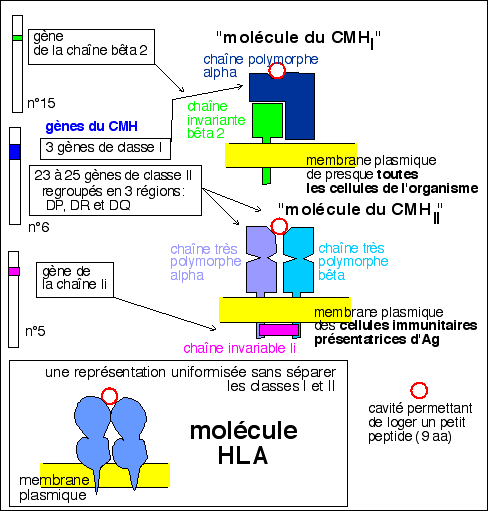
Une représentation TOTALEMENT THÉORIQUE des
"molécules du CMH"
(ces molécules ne sont pas
codées que par les gènes du CMH qui codent
par ailleurs pour de très nombreux autres
produits; de plus ces molécules sont des
glycoprotéines et pourtant les gènes qui
codent pour l'ajout des chaînes polysaccharidiques
(enzymes) ne sont pas évoqués; enfin il est
clair que s'il est indispensable de séparer les
deux classes de molécules)
Pour les élèves de terminale: une
représentation uniformisée de
"molécule HLA", que j'emploirais
désormais.
Remarque: les molécules du CMH (sans faire
de distinction de classe) ou molécules HLA sont
donc présentées en terminale comme des
marqueurs du soi qui sont
modifiés (soi
modifié) par la
présentation de déterminants
antigéniques. A un sujet du type: "vous
présenterez la notion de soi biologique et le
rôle de ses marqueurs dans la réponse
immunitaire" il faudrait traiter, sans
hésitation, les "molécules du CMH"
(différenciées ou non) et leur rôle
dans la réponse immunitaire.
Un antigène (Ag) est une molécule
déclenchant pas sa structure une réaction
immunitaire (reconnaissance par les
cellules immunitaires et réponse immunitaire
spécifique - voir plus loin). Un
déterminant antigénique
désigne plus précisément le
fragment de l'antigène qui a le rôle
antigénique.
Remarque: Lorsque l'on parle de
"présentation des déterminants
antigéniques en association avec le soi (les
molécules du CMH)" on fait ce me semble deux
erreurs: les petits peptides présentés avec
les "molécules du CMH" ne sont pas des
antigènes s'ils ne déclenchent pas de
réaction immunitaire. Et ensuite les
"molécules du CMH" ne sont pas des marqueurs du
soi (ni même du soi génétique), mais
par contre les petits peptides le sont en quelque
sorte.
|
Remarque concernant les greffes:
Les règles de sélection des organes à greffer
sont toujours un compromis entre les facteurs moléculaires
(histocompatibilité), l'état physiologique de l'organe
et surtout du donneur et l'urgence de la greffe. Pour toutes les
greffes on respecte la comptabilité ABO essentiellement pour
éviter de défavoriser les individus dont les groupes
sont les plus rares. Les incompatibilité au niveau des
produits des gènes du CMH NE SONT JAMAIS UNE CONTRE-INDICATION
à la TRANSPLANTATION d'organes. On observe cependant que le
pourcentage de survie des greffons à long terme augmente
légèrement en fonction du nombre d'identités A,
B (gènes codant pour la chaîne a
des molécules HLA de classe I)
et DR (région comportant de nombreux gènes
codant pour les deux chaînes polymorphiques des
molécules HLA de classe II) entre donneurs et receveurs. Les
greffes de moelle osseuse sont le plus souvent pratiquées
entre frères et sœurs génotypiquement HLA
identiques, sans tenir compte des groupes sanguins ABo.
Remarque importante:
Actuellement il semble qu'une vision très différente de
l'identité de l'organisme prédomine dans la
communauté scientifique : l'ensemble des molécules du
système immunitaire du groupe des anticorps formerait en
quelque sorte une image à la fois du monde
extérieur et du monde intérieur de l'organisme. Ces
concepts, très théoriques, ne sont heureusement pas au
programme.
2. l'homme, milieu de
vie
L'homme forme un volume qui n'est pas simple et dont les limites
ne sont pas facilement définissables. Classiquement on
distingue 3 volumes qui forment des milieux de vie différents:
le milieu extérieur (la peau étant la surface de
contact de l'homme avec le milieu extérieur), le milieu
extérieur internalisé (cavités internes en
relation directe avec le milieu extérieur par au moins un
orifice : bouche, tube digestif, vessie et canaux urinaires et
génitaux, utérus chez la femme, conduit auditif, fosses
nasales, trachée artère et poumons...; les surfaces de
contact sont humides (recouvertes de mucus) et moins
protégées que la peau) et le milieu
intérieur (avec des liquides internes: sang et lymphe).
Voici quelques éléments sur ces milieux,
préliminaires indispensables pour comprendre le lieu où
se déroule la réaction immunitaire.
2.1 Une surface séche peu franchissable : la
peau
La peau comprend deux couches d'inégales
épaisseur (vois schéma ci-dessous). La plus externe,
l'épiderme, comprend une couche de cellules de Malpighi (dont
la couche basale est formée de cellules souches qui se
divisent sans cesse et assurent le renouvellement complet de
l'épiderme en environ 20 jours) mais aussi des
mélanocytes (cellules pigmentaires), des cellules interveant
dans l'immunité (cellules de Langerhans) et des terminaisons
nerveuses libres. A la surface se l'épiderme on observe des
couches de cellules kératinisées (ou
kératinocytes, qui synthétisent et stockent à
l'intérieur de leur cytoplasme la protéine
(kératine) jusqu'à en mourir) : c'est la couche
cornée. Cet épiderme forme une barrière
qui limite avant tout les pertes d'eau par l'organisme (il se
dessécherait sinon car il vit en milieu aérien). Les
couches kératinisées sont effectivement
imperméables à l'eau mais pas aux gaz (la peau respire
mais l'homme rejette moins de 1% de son CO2 produit par respiration
par la peau (Schmidt-Nielsen, p 27)). Cette
barrière empêche aussi des corps étrangers de
pénétrer dans l'organisme. Mais comme toute
défense, elle a des points faibles : d'une part les
indispensables orifices d'échange (entre le milieu
intérieur et le milieu extérieur) et d'autre part sa
fragilité plus ou moins grande selon les zones
considérées.Vient ensuite le derme, irrigué et
innervé, comportant les organes sensitifs comme les
corpuscules tactiles de Pacini (voir cours de seconde), et des
glandes comme le follicule pileux sécréteur du poil ou
les glandes sudoripares (ou sudorales) ou encore les glandes
sébacées qui sont associées aux poils. La peau
repose sur des tissus sous-cutanés (hypoderme) contenant
notamment les couches de graisse (tissu adipeux formé de
cellules adipeuses ou adipocytes stockant les graisses).
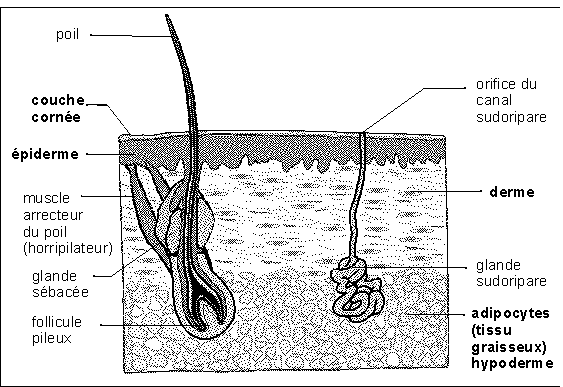
"Coupe schématique de peau humaine reposant sur une
couche de graisse" ou "lard d'homme"
(pour la petite histoire, on notera que la contraction du muscle
horripilateur (muscle lisse) provoque l'arrection (redressement) du
poil intervenant par exemple dans la chair de poule chez l'homme, ou,
plus efficacement, dans l'épaississement de la couche de
fourrure et donc d'un matelas d'air isolant chez les
mammifères à fourrure)
2.2 Une surface humide vulnérable : les
muqueuses
Une muqueuse est, du point de vue histologique
(l'histologie (du grec histos = tissu) est l'étude des
tissus), un épithélium (du grec epi = sur et
thêlé = mamelon) c'est-à-dire un tissu de
revêtement aux cellules adjacentes. Considéré du
point de vue fonctionnel c'est à la fois une barrière
et une zone d'échange... bref il intervient dans le travail de
relation de l'organisme. Une muqueuse est donc un
épithélium maintenu humide (elle
sécrète du mucus d'où son nom). Elle
revêt les organes en communication avec le milieu
extérieur, c'est vraiment l'interface "organisme - milieu
extérieur internalisé".
En voici un exemple, non pas pour le mémoriser mais pour
comprendre ce qu'est le lieu de vie de nombreux microorganismes et
donc un point de pénétration préférentiel
des germes pathogènes dans le milieu intérieur.
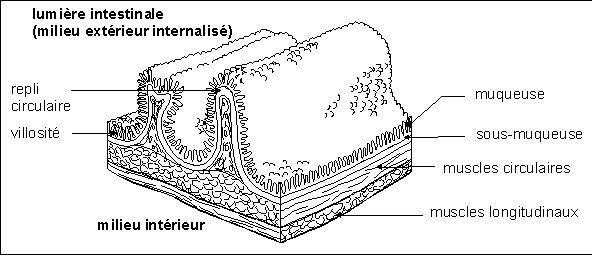
"Schéma simplifié de deux replis intestinaux" ou
"tripe d'homme"
(Pour se rendre compte de la surface d'échange
générée par le système de replis et
villosités, on peut citer le chiffre estimé d'environ
300 m2 pour les 4 à 6 mètres d'intestin chez
l'homme). La muqueuse comporte essentiellement les cellules
absorbantes (entérocytes) renouvelées en permanence (la
totalité de l'épithélium intestinal est
renouvelé tous les trois à cinq jours), les cellules
sécrétrices de mucus ainsi que d'autres cellules
sécrétrices aux rôles plus ou moins bien connus.
Dans la sous-muqueuse on trouve les vaisseaux sanguins très
nombreux ainsi que d'innombrables capillaires lymphatiques. C'est
aussi le tissu qui est innervé. La lumière intestinale
n'est pas un vide mais elle est comblée par les
matières en cours de digestion englobées dans les
sécrétions muqueuses, la "flore" intestinale... etc. La
quantité d'air (et surtout de gaz produit par les
réactions métaboliques fermentaires des
micro-organismes) est très réduite.
2.3 Des liquides internes très protégés :
sang et lymphe
Les tissus liquides de l'organisme sont essentiellement le sang et
la lymphe. Le sang vous est souvent plus familier mais la lymphe, vue
pourtant en classe de troisième théoriquement, demande
à être étudiée rapidement.
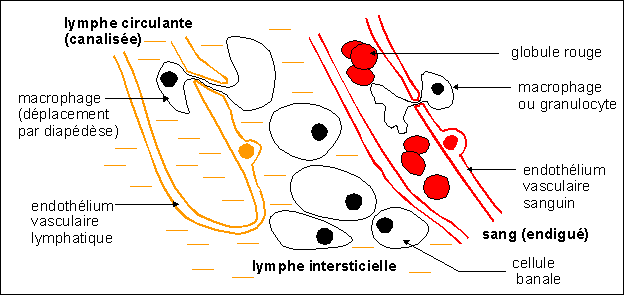
"Les trois compartiments liquidiens de l'homme" ou "le milieu
intérieur"
(sang - lymphe intersiticielle libre - lymphe circulante
canalisée). Le sang est un tissu (composé de cellules
réalisant une même fonction) dans un liquide (le
plasma : après coagulation, le plasma, dépourvu
d'une protéine : le fibrinogène, constitue le
sérum) qui est en mouvement: il circule grâce
à la pompe cardiaque et aux propriétés
élastiques des vaisseaux (voir programme de
spécialité). Sous pression, les liquides sanguins
filtrent à travers la paroi des capillaires sanguins (qui
retient les grosses protéines et bien sûr la plupart des
cellules) et vont constituer le liquide interticiel ou lymphe
intersticielle. Certaines cellules comme les macrophages sont
capables de passer entre les cellules de l'endothélium des
vaisseaux sanguins et passer dans les tissus. C'est le cas des
monocytes circulants dans le sang qui deviennent alors des
macrophages dans les tissus. De nombreux déchets comme le CO2,
diffusent à nouveau vers le sang et sont
récupérés par le système veineux, comme
la majorité des liquides extracellulaires. Mais une partie de
ces liquides, qui constituent la lymphe intersticielle, est
filtrée et récupérée dans les vaisseaux
lymphatiques qui draînent ainsi la plupart des tissus. La
lymphe canalisée venant de l'abdomen et des membres
inférieurs est draînée par le canal thoracique
puis se déverse passivement dans le sang au niveau du
carrefour veine jugulaire - veine sous-clavière gauche (voir
ci-dessous).
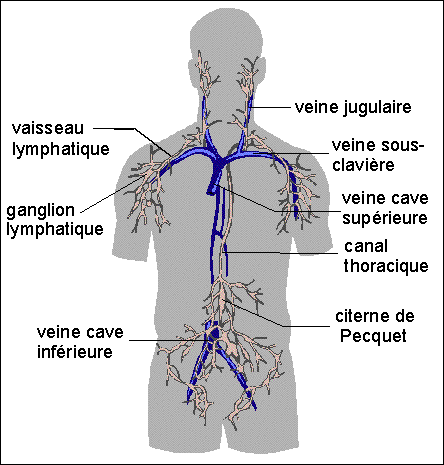
Système lymphatique de l'homme
(les vaisseaux sanguins veineux sont en bleu). Ni les vaisseaux
lymphatiques ni les ganglions ne sont contractiles. Les vaisseaux
lymphatiques, de même nature histologique que les vaisseaux
sanguins veineux, possédent cependant plus de valvules, qui
ont pour rôle, semble-t-il, d'assurer l'écoulement de la
lymphe dans un seul sens. La lymphe posséde essentiellement
des cellules immunitaires de type lymphocytes. Les ganglions
possédent un tissu lymphoïde essentiel à la
maturation des lymphocytes (leur structure et fonction n'est pas au
programme).
2.4. Quelques organismes qui "ont choisi" l'homme comme milieu
de vie
Quelques remarques préliminaires concernant quelques
relations entre les êtres vivants d'espèces
différentes (relations interspécifiques):
symbiose sens large
(s.l. ou sensu lato)
|
Etymologiquement (sym=avec et bio=vie), toute
association, ou communauté de vie, entre deux
êtres vivants est une symbiose. C'est le sens le plus
employé par les anglo-saxons à ma
connaissance. Il comprend dans ce cas le
commensalisme (du latin cum = avec et
mensa = la table), où un commensal tire
avantage d'un hôte qui n'est ni affecté, ni
aidé; le mutualisme (du latin mutuus =
réciproque) où le bénéfice est
réciproque pour les deux organismes associés;
et le parasitisme où l'hôte est
affecté par l'association avec le parasite.
On distingue classiquement les ectosymbiotes,
organismes symbiotiques qui restent à
l'extérieur de leur hôte (par exemple un poux
chez l'homme), et les endosymbiotes qui atteignent le
milieu intérieur (par exemple une bactérie
provoquant une infection).
|
symbiose sens strict
(s.s. ou sensu stricto)
ou mutualisme
encore symbiose vraie
|
Une symbiose (s.s.) est une association à
bénéfices réciproques.
|
|
parasitisme
|
Un parasite est un organisme qui vit de façon
obligatoire aux dépens d'un autre être vivant,
véritable milieu biologique. Le parasitisme est un
équilibre dynamique qui peut être rompu en
faveur du parasite (pathogénie) ou de l'hôte
(arrêt du développement du parasite pouvant
aller jusqu'à son élimination).
|
|
Remarque:
Les organismes ou les molécules à l'origine de
maladies (pathogènes) peuvent prendre plusieurs noms
: on a pris l'habitude de parler d'agent pour les
virus pathogènes (ou les viroïdes (acides
nucléiques nus) ou les prions (protéines
nues)... pour une discussion sur les caractères du
vivant voir la fiche sur la diversité
du vivant), de micro-organismes pour les
bactéries et les champignons, de parasites
pour les unicellulaires et les animaux. Cependant, on
emploiera, comme c'est l'usage, le terme très
général d'agent
pour désigner la cause directe d'une maladie
que celle-ci soit un agent (viral), un micro-organisme ou un
parasite.
|
De nombreux organismes vivent à la surface des enveloppes
externes et des muqueuses de l'homme. Savoir si ce sont des
symbiotes, au sens strict ou large, des parasites, ou s'ils ne sont
que des hôtes de passage, n'est pas facile et suppose toujours
une certaine interprétation, souvent discutable. Les cas de
pathogénie ne sont que des déséquilibres de
relations qui habituellement ne causent aucune maladie à
l'homme. Etant donné les innombrables espèces vivant
chez l'homme, en équilibre avec lui, les maladies sont en
quelque sorte les exceptions à la règle, l'insolite
déséquilibre... le désordre ?
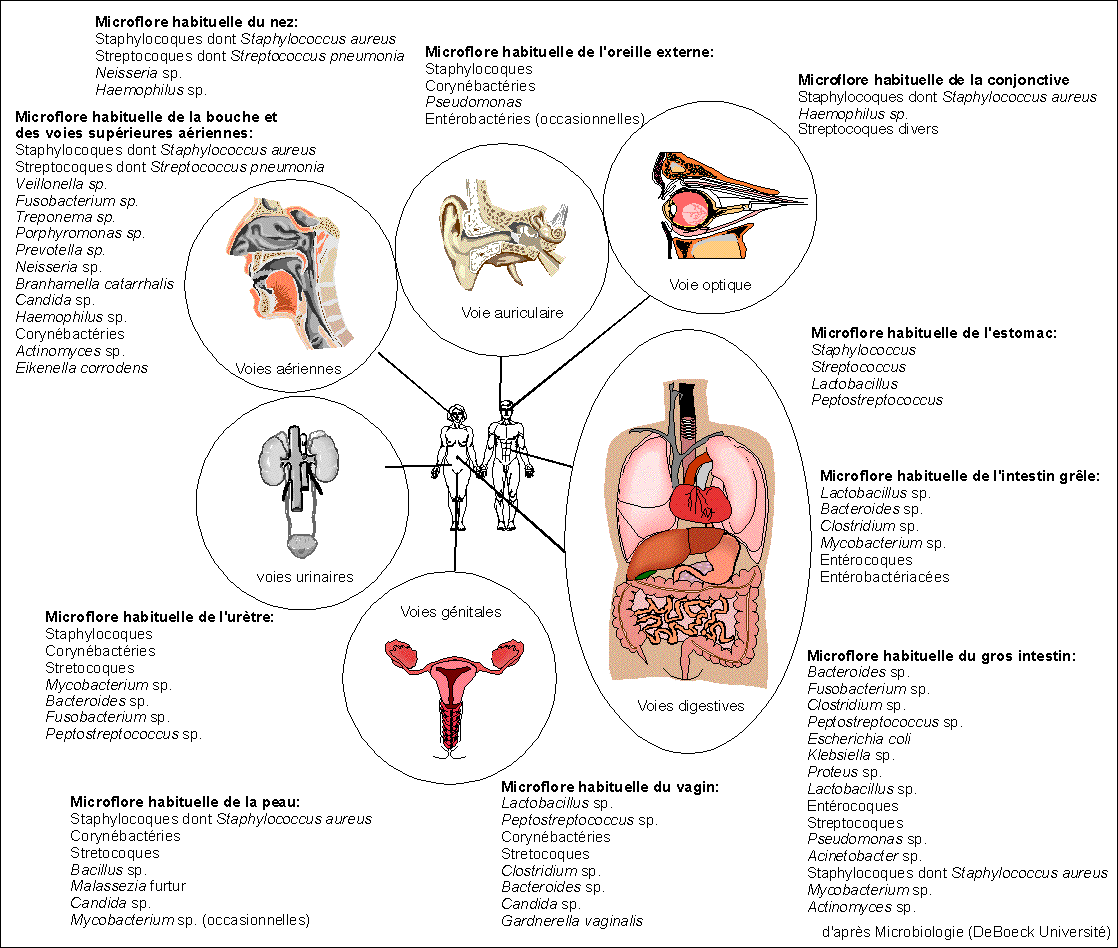
Quelques procaryotes formant la microflore habituelle de
l'homme
|
Exemples d'organismes eucaryotes vivant sur ou
chez l'homme
|
Nombreux
Protistes
|
Amibes, Coccidie (Tosxoplama gondi, agent de la
toxoplasmose), Plasmodium (agent du paludisme),
Ciliés, Trichomonas vaginalis (agent de vaginites et
d'urétrites : infections du vagin et de
l'urètre), Trypanosoma (agent de la maladie du
sommeil)....
|
Mycètes
(Champignons)
|
Pneumocystis carinii (agent de la pneumocystose
humaine)
|
|
Animaux
|
Schistosoma mansoni (agent de la bilharziose),
Ténia (ver solitaire), autres vers (Ascaris, Oxyure,
Filaires...), Poux, Tiques ...
|
Une étude intéressante concerne les
expériences d'élevages gnotobiotiques. C'est
Louis Pasteur qui, en 1897 semble avoir suggéré que les
animaux ne pouvaient se développer en l'absence de
micro-organismes. Des essais infructueux furent d'abord menés
sur des poulets axéniques (dont on détruisait la
microflore intestinale, du grec xéno= étranger
et "a" privatif) qui mouraient en moins d'un mois. Mais depuis
1912, on sait les élever avec un régime alimentaire
adéquat afin de remplacer le rôle des micro-organismes
intestinaux. On élève maintenant aussi des colonies de
rats, hamsters, lapins... axéniques en les prélevant
chez la mère et en les maintenant en conditions
stériles. On peut ensuite les inoculer avec telle ou telle
espèce de micro-organisme pour étudier son rôle :
on qualifie alors ces animaux d'élevage de
gnotobiotiques (du grec gnotos = connu et bio =
la vie). Les animaux axéniques n'ont pas une anatomie ni une
physiologie normales. Leur système immunitaire est par exemple
très réduit. Ils sont extrêmement sensibles aux
agents pathogènes.
On peut résumer en termes de barrières les
mécanismes de défense de l'hôte intervenant avant
que l'organisme étranger supposé pathogène n'ait
pénétré à l'intérieur de
l'organisme humain (dans le milieu intérieur).
|
Les barrières
s'opposant chez l'homme à la
pénétration d'un organisme
étranger
|
|
barrières biologiques
|
l'état de nutrition
|
un individu mal nourri est plus sensible aux maladies
qu'un individu bien nourri...
|
|
l'âge.
|
en général les individus très jeunes
et très vieux sont les plus vulnérables...
|
|
les microflores habituelles
|
la "flore" habituelle (indigène normale)
entrant en compétition (pour l'espace et
pour les éléments nutritifs) avec un organisme
pathogène étranger, elle peut aussi produire
des substances antibactériennes toxiques pour les
autres espèces
|
|
terrain "héréditaire"
|
les variations individuelles sont indiscutables. Leur
cause est par contre fort discutée... je vous propose
de vous reporter à ce que l'on a vu dans la
première partie sur la connaissance du soi et de son
hérédité.
|
|
barrières physico-chimiques
|
la peau
|
la couche de cellules kératinisées est
difficile à percer et se renouvelle sans cesse, la
peau est sèche, recouverte du sebum acide
secrété par les glandes
sébacées, la peau est déjà
occupée par de nombreux organisme habituels, l'homme
enfin se lave (barrière
comportementale)...
|
|
le mucus
|
le mucus collant et les cils des muqueuses (voir plus
haut) agglutine les organismes étrangers, la toux et
l'éternuement peuvent aussi dégager
brusquement les voies respiratoires...
|
|
les écoulements de fluides
|
larmes de l'oeil et mictions (action d'uriner), par
exemple, éliminent de nombreux micro-organismes.
|
|
le pH
|
pH gastrique voisin de 2-3
|
|
les antimicrobiens naturels
|
de nombreuses substances sont des antimicrobiens : comme
le lysozyme des larmes ou certaines enzymes
salivaires ou encore les interférons (famille de
glycoprotéines produites par de nombreuses cellules
eucaryotes en réponse à divers stimuli comme
des infactions vrales...)
|
1 . A la recherche de l'identité de l'homme ;
2 . L'homme, milieu de vie ;
3 . L'homme blessé ; 4 . L'homme
malade; 5 . L'homme sain
TP: le système immunitaire: des
cellules et des organes -
TD: retour sur les cellules et les
molécules du système immunitaire
retour sommaire cours Terminale
S