Réflexes myotatiques et
réflexes de flexion
Sources
Encyclopedia universalis (tonus
musculaire, motricité)
Précis de physiologie, Calas et al., 1997, Doin
Le réflexe myotatique est le seul réflexe
monosynaptique connu. Le tonus musculaire se
définit comme la résistance d'un muscle à son
étirement. Il est assuré par des réflexes
myotatiques.
Les réflexes de flexion sont polysynaptiques.
Ce sont des réflexes de défense impliquant le retrait
d'un membre à la suite d'un stimulus nociceptif
1 . Le tonus musculaire, résistance d'un muscle à
son étirement, met en jeu des réflexes myotatiques,
seuls réflexes monosynaptiques connus.
Dans l'étude de la motricité, on a opposé
historiquement la contraction tonique d'un muscle
squelettique, soutenue, permanente, parfois qualifiée de
statique car elle intervient d'abord dans la posture, à
la contraction phasique, volontaire et brusque mais
temporaire, parfois qualifiée de dynamique car elle
entraîne le mouvement. Actuellement on pense que ces deux
composantes existent pour tous les muscles et sont étroitement
liées. Différents types de fibres musculaires
(plus ou moins fatigables et rapides)
sont aussi reconnus histologiquement. Deux principes essentiels ont
été dégagés des études
histologiques et embryologiques:
* chaque unité motrice (groupe de fibres
innervées par un seul motoneurone) est composée de
fibres homogènes
* c'est le motoneurone qui, au cours du développement de
l'organisme, détermine le type des fibres musculaires qu'il
innerve; On sait expérimentalement modifier le type des fibres
musculaires en provoquant l'innervation d'une unité motrice
par un autre motoneurone que celui naturellement mis en place.
Le tonus musculaire est à la fois un état de
tension légère et la résistance à
l'étirement d'un muscle squelettique sain in
vivo. Le tonus musculaire est d'une extrême
importance en ce qu'il constitue le point de départ des
activités motrices et posturales, préparant le
mouvement, fixant l'attitude, sous-tendant le geste, maintenant la
statique et l'équilibre. Son importance va de pair avec une
complexité due à l'intervention de l'ensemble des
structures nerveuses centrales.
|
Réflexe d'étirement:
Quand un muscle est étiré les
fuseaux neuromusculaires sont
aussi étirés et leurs
fibres sensorielles
(Ia
myélinisées (d'assez gros
diamètre : 12 à 18 µm) et II
(de diamètre plus petit: 6 à
10 µm et non représentées
ci-contre) envoient des PA dont la fréquence
est proportionnelle à l'allongement (codage statique)
et à la vitesse de l'élongation (codage
dynamique). En réponse réflexe l'ensemble du
muscle étiré se contracte par envoi d'un
message moteur par l'intermédiaire des
motoneurones alpha. Une
stimulation des neurones sensoriels en T des ganglions
spinaux dont les dendrites sont reliés aux fuseaux
neuromusculaires provoque la même réponse. Le
réflexe d'étirement est donc un réflexe
qui s'oppose à l'étirement involontaire d'un
muscle. C'est ce que l'on appelle le tonus
musculaire. Plus un muscle a du tonus plus il est
capable de résister à son étirement. Un
muscle sans tonus s'étirant sans opposer de
résistance ne serait pas de grande utilité
dans le maintien de la posture.
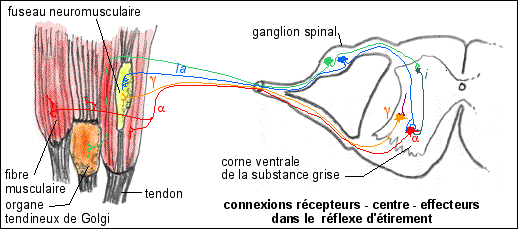
Un réflexe d'étirement
d'après Précis de physiologie, Calas et al.,
1997, Doin, fig 10.18 p 341
|
|
Mais avec les seuls fibres
Ia et
alpha le muscle ne pourrait
avoir qu'une longueur fixe.
Lorsque l'on stimule directement les motoneurones alpha le
muscle se contracte sans que les fuseaux neuromusculaires
soient étirés. Le relâchement des fibres
fusoriales provoque même un arrêt des PA
sensitifs envoyés aux ganglions spinaux. Pour
éviter cette perte d'information et ajuster la
longueur du muscle à une valeur non figée les
fibres du fuseau neuromusculaire peuvent aussi se contracter
sous l'effet de la stimulation de
motoneurones gamma qui
innervent les fibres fusoriales (axones
myélinisés de petit diamètre: 4
à 8 µm). Ainsi lors d'une contraction
volontaire (par voie
descendante venant des différentes aires
cérébrales) les fibres fusoriales se
contractent en même temps que les fibres musculaires
banales du muscle par stimulation des motoneurones gamma.
Mieux encore c'est par l'intermédiaire de ces
motoneurones gamma que la contraction volontaire d'un muscle
est déclenchée. En effet, si la contraction
des fibres fusoriales est forte alors que les fibres
musculaires banales sont relâchées, le nombre
de fibres fusoriales est insuffisant pour provoquer une
contraction puissant du mus3cle mais un réflexe
d'étirement est déclenché et c'est par
l'intermédiaire des neurones sensitifs (fibres Ia et
II) que les motoneurones alpha sont activés et qu'ils
provoquent une contraction puissante et efficace. C'est
un étirement volontaire à travers le
réflexe d'étirement; c'est ce que l'on appelle
la "boucle gamma".
D'une façon encore plus précise il semble
qu'il y ait une permanente adaptation de la longueur du
fuseau musculaire à l'état d'élongation
du muscle lors de la contraction volontaire, ce qui permet
à ce mécanorécepteur de rester sensible
à l'étirement tout au long de la
contraction.
|
|
Lors d'un réflexe d'étirement et pour qu'il
y ait mouvement réflexe, les muscles antagonistes des
muscles étirés qui se contractent doivent
aussi se relâcher. Cette inhibition des muscles
antagonistes serait due à une innervation
réciproque à partir des motoneurones alpha.
Les fibres sensitives en liaison avec les fuseaux
neuromusculaires établiraient une liaison directe
avec les motoneurones alpha contrôlant la contraction
du muscle étiré et une liaison indirecte par
l'intermédiaire d'interneurones inhibiteurs avec les
motoneurones innervant les muscles antagonistes.
|
|
Le fuseau neuromusculaire,
récepteur à l'étirement comprend 2
à 10 fibres musculaires modifiées (peu
striées) encapsulées (capsule conjonctive)
attachées aux membranes complexes
(aponévroses) cloisonnant le muscle. Deux types de
fibres fusoriales se rencontrent: 2 à 3 fibres
à "bague nucléaire" dont la partie centrale
est renflée et 3 à 5 fibres "à
chaîne nucléaire" présentant une zone
allongée où sont condensés les
noyaux.
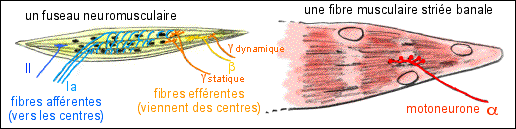
D'après Précis de
physiologie, Calas et al., 1997, Doin, fig 10.19 p 341
|
Les contrôles médullaires:
* Les organes neurotendineux de Golgi
sont sensibles avec un seuil assez bas à la contraction de
l'ensemble du muscle mais sont peu sensibles à son
étirement. En cas de contraction volontaire ou réflexe
ils peuvent limiter la contraction en agissant sur les motoneurones
alpha par l'intermédiaire d'un
interneurone inhibiteur
(i nommés "neurones
en bouteille" de Golgi). Ils peuvent éviter ainsi des
déchirements tendineux.
Un muscle hypertonique dont l'activité gamma
est très élevé peut présenter ce que l'on
appelle la réaction de la "roue dentée". Soumis
à une flexion passive du coude, la résistance à
l'étirement augmente puis s'effondre brusquement.
D'autres récepteurs intraligamentaires, cutanés,
articulaires..., sont aussi en relation avec les motoneurones au
niveau médullaire mais par des voies encore mal connues.
* L'inhibition récurrente des motoneurones.
Les motoneurones alpha (notés
alpha f) ont des collatérales qui sont en relation avec des
interneurones inhibiteurs appelés
cellules de Renshaw eux même en relation avec les
motoneurones. Une stimulation excessive des motoneurones (par la
voie descendante notamment) est ainsi
amortie par inhibition du motoneurone stimulé par les cellules
de Renshaw en contact avec lui-même. Mais les cellules de
Renshaw sont aussi en relation, par l'intermédiaire
d'interneurones inhibiteurs, avec les
motoneurones alpha (notés alpha
e) innervant les muscles antagonistes, qu'ils stimulent donc
indirectement.
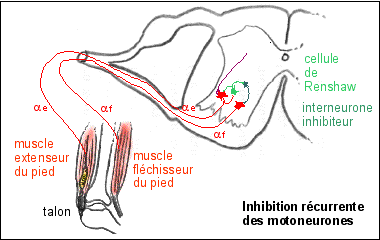
D'après Précis de physiologie, Calas
et al., 1997, Doin, fig 10.22 p 345
Le neurotransmetteur des motoneurones alpha est
l'acétylcholine, aussi bien au niveau des fibres
musculaires que des cellules de Renshaw. Les cellules de Renshaw
sécrètent de la glycine, un neurotransmetteur
inhibiteur dont les récepteurs synaptiques peuvent être
bloqués par la strychnine (antagoniste).
L'empoisonnement à la strychnine conduit à des
tétanies (du grec "tenein" = tendre;
contractions maximales prolongées observées
historiquement lors des phases terminales de l'infection par le
bacille tétanique) et des convulsions.
L'importance de ces afférences centrales peut
être mise en évidence lors de la section des racines
médullaires dorsales. Le muscle innervé est alors a
demi-paralysé alors que son innervation motrice est
intacte.
Des contrôles centraux:
D'innombrables stimulations centrales sont susceptibles de
modifier l'activité de base des motoneurones gamma et parfois
directement des alpha. On peut citer la rotation de la tête
(stimulation des récepteurs cervicaux et des récepteurs
de l'oreille interne), la stimulation du cervelet ainsi que diverses
structures du mésencéphale (noyau rouge en
particulier), du diencéphale (certains noyaux du thalamus
dorsal) et du télencéphale (corps striés et
cortex cérébral lui-même) ou encore le tronc
cérébral.
2. Les réflexes de flexion
Historiquement les réflexes de flexion ont
été mis en évidence chez la grenouille et sont
connus sous le nom de lois de Pflüger:
- loi de localisation: après une stimulation douloureuse de la
peau d'une patte on observe la contraction des fléchisseurs
qui retirent la patte stimulée;
- loi de symétrie: pour une intensité supérieure
du stimulus, la flexion de la patte stimulée s'accompagne de
celle de la patte symétrique
- loi d'irradiation: la réponse se transmet aux membres
opposés antérieurement ou postérieurement;
- loi de généralisation; la réponse se transmet
à toute la musculature.
Chez le Chien ou la chat spinal (dont la moelle
épinière est sectionnée) la réponse de la
patte symétrique est une extension.
Ces réflexes existent aussi chez l'homme. Après une
irritation de la plante du pied, on observe habituellement une
flexion des orteils. Chez l'homme spinal ou ayant subi certaines
attaques cérébrales, on observe au contraire une
extension du gros orteil ainsi que des autres orteils en
éventail (signe de Babinski).
Ces réflexes sont des réflexes de défense
et polysynaptiques (le temps de réponse et long et la
réponse est prolongée, même après
arrêt de la stimulation). Ils sont considérés
comme le reflet de comportements stéréotypés
spécifiques d'un organisme liés à son mode de
vie et à son type de locomotion notamment.